丝状真菌瑞氏木霉生产重组蛋白的分子生物学研究进展(一)
- 格式:docx
- 大小:23.91 KB
- 文档页数:9


微生物学通报 MAY 20, 2010, 37(5): 738−747 Microbiology China © 2010 by Institute of Microbiology, CAStongbao@基金项目:国家863计划项目(No. 2006AA10A211) *通讯作者:Tel: 86-531-82605386; Fax: 86-531-82965636; : yanght@收稿日期:2009-08-24; 接受日期:2010-01-18专论与综述植酸酶的多样性及其分类李晓龙1,2 杨合同1,2* 扈进冬2 吴远征2 李纪顺2 任艳2(1. 山东理工大学生命科学学院 山东 淄博 255049)(2. 山东省科学院生物技术中心 山东省应用微生物重点实验室 山东 济南 250014)摘 要: 植酸酶是一类催化植酸水解逐步释放磷酸基团形成低级肌醇磷酸衍生物的正磷酸单酯磷酸水解酶。
植酸酶在动物营养、资源环境保护和人类健康等领域有巨大的应用潜力。
目前, 人们对植酸酶的多样性及其分类的认识比较模糊甚至错误, 严重影响了植酸酶的研究进程和水平。
首先简要概述了基于最适pH 和立体专一性的植酸酶分类, 然后着重论述了基于结构和催化机理的植酸酶分类及其代表酶特征的最新研究进展, 最后探讨了根据不同分类标准特别是基于结构和催化机理准确理解和全面表征各种植酸酶的重要性, 以期为植酸酶的研究和应用提供参考。
关键词: 植酸酶, 最适pH, 立体专一性, 结构和催化机理, 分类Diversity and Classification of PhytasesLI Xiao-Long 1,2 YANG He-Tong 1,2* HU Jin-Dong 2 WU Yuan-Zheng 2LI Ji-Shun 2 REN Yan 2(1. School of Life Sciences , Shandong University of Technology , Zibo , Shandong 255049, China )(2. Shandong Provincial Key Laboratory for Applied Microorganism , Biotechnology Center , Shandong Academyof Sciences , Jinan , Shandong 250014, China )Abstract: Phytase is a type of orthophosphomonoester phosphohydrolase, which catalyticly initiates step-wise hydrolysis of phytic acid to release phosphate radicals and produce lower inositol phosphate deriva-tives. Phytase has great application potential in the areas of animal nutrition, resource conservation, envi-ronmental protection and public health. At present, the understanding in phytase diversity and classification is so confusing or even inaccurate that the research progress and level of phytase has been badly affected. In this paper, the phytase classification based on optimal pH and stereospecificity is briefly introduced first, and then the updated research advance in phytase classification based on structure and catalytic mechanisms and the attributes of the representative phytases are summarized and discussed comprehensively. It is of vital importance to take into consideration the classification standards especially to focus on structure and cata-lytic mechanisms when a given phytase could be fully and accurately understood and characterized. Keywords: Phytase, Optimal pH, Stereospecificity, Structure and catalytic mechanisms, Classification 植酸(Phytic acid 或Phytate), 即肌醇六磷酸(myo -Inositol hexa kis phosphate, IP6), 是植物种子中磷元素的主要储存形式, 普遍存在于植物性食品和饲料[1]。

《利用重组蛋白技术提高Taq DNA聚合酶活性的研究》篇一一、引言随着生物科技的迅速发展,重组蛋白技术在生物学研究及实际应用中发挥着越来越重要的作用。
Taq DNA聚合酶作为分子生物学领域中常用的酶类,其活性直接影响到PCR(聚合酶链式反应)的效率和准确性。
因此,提高Taq DNA聚合酶的活性成为了一个重要的研究方向。
本文旨在探讨利用重组蛋白技术提高Taq DNA聚合酶活性的研究。
二、材料与方法1. 材料本研究所用材料主要包括Taq DNA聚合酶基因、表达载体、宿主细胞以及相关的实验试剂。
2. 方法(1)基因克隆:首先,从已知的DNA序列中获取Taq DNA 聚合酶的基因序列,然后将其克隆到表达载体中。
(2)重组蛋白表达:将含有Taq DNA聚合酶基因的表达载体转入宿主细胞中,通过诱导表达获得重组Taq DNA聚合酶。
(3)酶活性检测:通过PCR反应检测重组Taq DNA聚合酶的活性,并与其天然酶活性进行比较。
(4)优化表达条件:通过改变表达条件(如温度、pH值、诱导剂浓度等),优化重组Taq DNA聚合酶的活性。
三、实验结果1. 重组Taq DNA聚合酶的表达通过基因克隆和重组蛋白表达,成功获得了重组Taq DNA聚合酶。
SDS-PAGE结果表明,重组Taq DNA聚合酶在宿主细胞中得到了有效表达。
2. 酶活性检测PCR反应结果显示,重组Taq DNA聚合酶具有较高的活性。
与天然Taq DNA聚合酶相比,重组Taq DNA聚合酶的活性得到了显著提高。
3. 优化表达条件通过改变表达条件,发现适当的温度、pH值和诱导剂浓度能够进一步提高重组Taq DNA聚合酶的活性。
其中,在37℃、pH 值为7.5的条件下,使用适量的诱导剂,能够获得最佳的酶活性。
四、讨论本研究利用重组蛋白技术成功提高了Taq DNA聚合酶的活性。
这为PCR反应的效率和准确性提供了有力保障,有助于推动分子生物学领域的发展。
此外,通过优化表达条件,可以进一步提高重组Taq DNA聚合酶的活性,为实际应用提供了更多可能性。

万方数据万方数据·140·董方,等.平分型G1cNAc糖基化修饰的生物学功能蛋白分选的负性调节信号n21。
3.3生殖发育调控人类精子缺乏MHCI类分子,使其易于被NK细胞溶解。
而MHcI阴性的肿瘤细胞通过在肿瘤细胞表面表达足量的平分型N一聚糖,而阻断NK细胞的溶解作用。
因此,精细胞也可能通过同样的机翩选避NK的细胞溶解作用。
利用超敏质谱仪对精细胞N一聚糖测序可以检测到3类N一聚糖:①高甘露糖塑;②二天线平分型;③以hisx和IJewis丫作为终端修饰的二天线、三天线、四天线型。
糖蛋白染色显示,携带呐穿列糖蛋白分布于顶体莉不是位于浆膜;丽功能缺陷的精子则表现为抗娜抗体分布异常(图2)。
推测惨饰聚糖可以抑制生殖道内计对精子糖蛋白的抗原特异性免疫反嘘。
嘲』III匕,人类精子哟主要聚糖与先天性及获得性免疫反瘟哟抑制有关。
这些结果支持哺乳类动物表邀碳水化合物与|配子《耨删魄哆保护及人类在子宫棚哟发育有关。
某些特殊类型的细胞表达特异性糖姐模式,惜此发挥相嘘睡句生物学功能‘引。
嘲2久类储子表达的主要N-聚糖k:高甘露糖型(Ma呜9tol湖舢:);瞻::件有或不件宥核心者澡糖者藻糖修饰哟平分型二天线结构;lO:盖、兰、嘲毙线聚糖,板心砻澡糖岩藻糖修饰,镌带呱嫡毋致/或蜥r谬饰赃娠lf目关糖蜚囟【(畔e粤l堋呀啪舯幽峨硼挈蜘o_删坩i拣,‰9是反刍勘物滋养层细胞分泌韵主要蛋白。
,遨些蛋白储存在滋养屡巨大细胞分秘颗粒恂.,与母体子宫止发细胞融合后释放进入母体器。
雷。
妊娠串期Ip黻最丰寓的弘聚糖是四无线核心绪!藻糖基眈修饰辅构’,并连接有平分型蛾Nk。
组成湖8航塬[№№娆.,3(·G姗№鲫、49l创必.,删l渊№]哟聚糖均为删天线型。
采用识别铷薯睨原的单克糟醐谢:(心埴ll9对分泌颗粒染色,分断:显示、,鳃娠.娩出话歪分娩前数天.,可以检测判黜a航原的存在。
I山褴凝集素t(t脚溅i珏删脚I卜sislectin)(该凝集素可以与a2,3·唾液酸修饰的乳糖氨)组织化学分析显示,该分泌颗粒在妊娠32d及分娩前呈阳性。



漆酶基因异源表达及其酶活性的研究进展刘晓庆;那日;郭九峰【摘要】近年来由于漆酶在生物漂白和农作物秸秆利用等方面具有广阔的应用前景,对漆酶的研究越来越受到国内外学者的重视.然而,自然界漆酶的产量和酶活较低,难以适应工业化生产需求.此外,漆酶的产酶效率低,成本高.实现漆酶的异源高效表达是解决这一问题的有效途径.目前,国内外已有多种来源的漆酶基因被成功克隆,并在不同的宿主细胞中实现异源表达,但迄今为止漆酶基因异源袁达结果仍然不理想,离真正实现漆酶的高效表达还有一定距离.【期刊名称】《安徽农业科学》【年(卷),期】2016(000)001【总页数】4页(P18-21)【关键词】漆酶;宿主细胞;异源表达;基因克隆;酶活性【作者】刘晓庆;那日;郭九峰【作者单位】内蒙古大学物理科学与技术学院,内蒙古呼和浩特010021;内蒙古大学自治区离子束生物工程重点实验室,内蒙古呼和浩特010021;内蒙古大学自治区离子束生物工程重点实验室,内蒙古呼和浩特010021;内蒙古大学自治区离子束生物工程重点实验室,内蒙古呼和浩特010021【正文语种】中文【中图分类】S188漆酶(laccase,Ec1.10.32)属于含铜的多酚氧化酶,能有效降解自然界中木质素等复杂有机物。
真菌和植物中都可以分泌漆酶,少数的昆虫和细菌也可以分泌漆酶,而真菌是漆酶的主要生产者。
它通过获得O2对苯酚物质进行催化作用,而生成物中仅有水。
漆酶是一种不可多得的环保型氧化酶。
漆酶是参与自然界木质素循环与利用的重要酶之一。
漆酶对富含木质素的农作物残渣的有效降解作用不仅可以减少秸秆焚烧带来的环境问题,而且可以提高青贮饲料的品质和利用率,对与木质素结构相似的污染物也有明显的降解能力。
据统计,漆酶有作用底物250多种[1]1。
漆酶的底物多样性决定了它是一种多功能氧化酶。
它在工业、农业及食品工业都有着极其广泛的应用。
近年来研究表明,漆酶在生物技术方面也有重要的作用,可用于生物传感器与生物检测。

第28卷第4期江西农业大学学报Vol.28,No.4 2006年8月Acta Agriculturae Universitatis J iangxiensis Aug.,2006文章编号:1000-2286(2006)04-0516-05丝状真菌遗传转化系统研究进展李娟,杨金奎,梁连铭,张克勤3(云南大学生物资源保护与利用重点实验室,云南昆明650091)摘要:近年来,丝状真菌的分子生物学研究发展非常迅速,尤其是丝状真菌遗传转化研究取得了长足的进展。
对丝状真菌遗传转化系统的最新研究进展进行综述,主要包括丝状真菌的转化方法、选择标记、转化系统的应用等。
关键词:丝状真菌;转化方法;选择标记中图分类号:Q949.32 文献标识码:AAdvance i n the Research on Geneti c Transfor mati onSyste m of Fil amentous FungiL I Juan,Y ANG J in-kui,L I A NG L ian-m ing,ZHANG Ke-qin3(Laborat ory for Conservati on and U tilizati on of B i o-res ources,Yunnan University,Kunm ing650091,China) Abstract:Fila ment ous fungi are a kind of eukaryoteswhich exist universally in nature and are of econom i2 cal significance.The latest p r ogresses in the research on genetic transf or mati on syste m p lay an i m portant r ole in understanding fila ment ous fungi.So me transf or mati on methods,selective markers,and the app licati on of transf or mati on syste m in the field of research on fila ment ous fungi are revie wed in this paper.Key words:fila ment ous fungi;transf or mati on method;selective marker1 概 述丝状真菌作为低等的真核生物,对人类生命活动的各个方面均产生了深远的影响,为了控制真菌对人类不利的一面,充分利用其有益的特性服务于人类,就必须深入的了解其生理、遗传以及分子生物学方面的信息,并通过分子生物学手段对其进行有效改造。


第24卷第12期2012年12月生命科学Chinese Bulletin of Life SciencesV ol. 24, No. 12Dec., 2012文章编号:1004-0374(2012)12-1384-59国家自然科学基金委员会生命科学部2012年度面上项目项目名称申请人依托单位1微生物学渤海海滨湿地螺旋藻、节旋藻资源调查及基于DNA序列分析的分类系统重建王素英天津商业大学铁皮石斛内生菌多样性及其促进铁皮石斛活性成分积累的机理研究刘文洪浙江中医药大学锦鸡儿属根瘤菌生态适应性和分子进化研究陈文峰中国农业大学我国部分城市环境水系中军团菌分离株遗传多态性研究宋亚军中国人民解放军军事医学科学院鸭绿江滨海湿地放线菌多样性及农用活性化合物资源的发掘于基成大连民族学院菊三七属药用植物内生放线菌多样性及难培养物种的分离张秀敏河北大学泛基因组学指导下的拟无枝酸菌属分子分类学模型构建及重要次生代谢郑华军上海人类基因组研究中心产物挖掘八种珍稀动物粪便放线菌多样性研究姜 怡云南大学中度嗜盐拟诺卡氏菌嗜盐机理研究李文均云南大学嗜盐放线菌功能基因及其次生代谢产物资源的挖掘唐蜀昆云南大学斑痣盘菌目新增地锤菌科分类学与二科系统高海拔地区物种多样性研究林英任安徽农业大学腐霉属的DNA条形码研究韦继光广西大学粒毛盘菌属黑色素活性筛选、分子修饰及构效关系研究叶 明合肥工业大学中国丝盖伞属和相近属的分类及分子系统学研究图力古尔吉林农业大学拟康氏木霉合成的康宁霉素促进植物根系生长的作用与机制宋晓妍山东大学中国热带及亚热带地区文字衣属地衣分类学及其生物地理学研究贾泽峰山东农业大学中国小网衣及其相关属地衣的系统分类学研究王海英山东师范大学中国块菌属分类学及分子系统学研究范 黎首都师范大学西藏冬虫夏草遗传多样性及与虫草品质相关性研究旺 姆西藏农牧学院甘薯长喙壳菌在单双子叶植物间相互侵染的机制研究黄 琼云南农业大学紫色红曲菌色素和桔霉素合成关键基因的克隆及表达调控蒋冬花浙江师范大学白环蘑属(蘑菇科)真菌的分类及分子系统发育研究葛再伟中国科学院昆明植物研究所中国乳牛肝菌属及其两个重要复合群的系统学和生物地理学研究刘培贵中国科学院昆明植物研究所藏东川西横断山地区黑粉菌锈菌物种多样性及区系特征郭 林中国科学院微生物研究所Euops属卷叶象甲与共生真菌的协同进化及化学机制刘杏忠中国科学院微生物研究所石耳目科属共祖基因序列及表型综合性状的研究魏江春中国科学院微生物研究所丝膜菌科系统学研究魏铁铮中国科学院微生物研究所中国肉座菌科分类以及该科DNA条形码和分子系统学研究庄文颖中国科学院微生物研究所广义申克孢子丝菌的分子系统发育与生理病理特征分化研究周 汛重庆医科大学温度影响新型极端微生物Anoxybacillus flavithermus subsp. yunnanensis有机溶剂彭 惠安徽大学耐受性的机理研究具有抗肺癌作用的三株真菌次生代谢产物及作用机理的研究刘为忠滨州医学院细菌性软腐病菌Cpx双组分信号转导系统的功能及其调控机制周晓辉河北科技大学蓝细菌集胞藻6803中S2P蛋白酶介导级联信号转导参与胁迫响应的机制研究陈 谷华南理工大学基于色素蛋白复合体的DSSC的新型宽光谱高效染料的筛选与合成杨素萍华侨大学c-di-AMP候选受体蛋白TerD的结构功能及其信号途径研究何 进华中农业大学“桑黄”类药用菌的次生代谢产物资源及其部分化合物的抗肿瘤活性机理研究包海鹰吉林农业大学∙ 基金∙第12期1385国家自然科学基金委员会生命科学部2012年度资助项目光滑球拟酵母能量代谢调控其环境适应性的生理机制刘立明江南大学Candida glycerinogenes过量合成甘油过程中渗透压对EMP/HMP途径代谢偏转诸葛斌江南大学的调控研究Pseudomonas putida DLL-E4对硝基苯酚降解中的温敏与转录调控机制崔中利南京农业大学毕赤酵母多不饱和脂肪酸生物合成相关脱氢酶基因的功能及其分子调控机制李明春南开大学的研究微生物乳酸代谢调控蛋白LldR的结构及机制研究马翠卿山东大学组学工具研究拟威克酵母变种的槐糖脂合成酶系及调控机制宋 欣山东大学蓝细菌Synechocystis sp. PCC 6803强光诱导多肽功能及强光适应分子机制何庆芳山东省农业科学院的研究群体感应系统阻遏抗真菌剂藤黄绿菌素生物合成的分子机制黄显清上海交通大学安莎类抗生素聚酮链释放与环化机制的结构生物学研究康前进上海交通大学绿针假单胞菌GP72中吩嗪类化合物合成的正向自调控机制研究张雪洪上海交通大学双组分系统在普通脱硫弧菌金属腐蚀中的调控机制陈 磊天津大学基于微环境调控的产多杀菌素刺糖多孢菌代谢机制研究卢文玉天津大学天然木糖利用麦芽糖假丝酵母葡萄糖阻遏效应及其调控机理研究王钦宏天津工业生物技术研究所基于新型胸腺嘧啶补偿途径的多氧霉素组分多样化的分子机制研究陈文青武汉大学微生物药物生物合成转化知识库的构建胡黔楠武汉大学基于嗜热、嗜温及嗜冷菌来源的三种高度同源蛋白酶的温度适应性分子唐 兵武汉大学机制研究假单胞菌NyZ12降解环己胺的分子机理研究闫达中武汉工业学院谷氨酸棒杆菌对木质纤维素糖化液中酚类抑制物质的耐受机理研究沈锡辉西北农林科技大学狭叶南五味子内生真菌的代谢产物和功能研究董锦艳西南大学球孢白僵菌锌指蛋白基因Bbazf在卵孢素合成中的功能范艳华西南大学希瓦氏菌硝酸还原多层次调控的分子机制高海春浙江大学海洋动物寄生白僵菌Beauveria felina EN-135代谢产物分子多样性与活性研究王斌贵中国科学院海洋研究所Synechocystis PCC 6803缺铁诱导蛋白IsiA相互作用网络的构建及其功能研究王 强中国科学院水生生物研究所瘤胃细菌H1纤维素酶合成的群感效应调控研究东秀珠中国科学院微生物研究所丙酮丁醇发酵过程中菌体自溶现象的遗传学基础研究李 寅中国科学院微生物研究所禾粟链霉菌谷氏菌素生物合成基因簇的功能研究谭华荣中国科学院微生物研究所凝结芽孢杆菌产高光学纯L-乳酸的机理研究王丽敏中国科学院微生物研究所细菌分解代谢2-氯-4-硝基酚污染物的机理研究张俊杰中国科学院武汉病毒研究所铁、氧代谢途径的关键基因在控制趋磁螺菌磁小体合成中的作用李 颖中国农业大学苏云金芽胞杆菌Sigma 54因子调控的代谢途径与Cry蛋白产量的关系宋福平中国农业科学院植物保护研究所Rad53磷酸化模式在白色念珠菌细胞周期检验点及形态发生调控中的作用机制桑建利北京师范大学原核生物多功能蛋白trigger factor体内生理作用机制的研究刘川鹏哈尔滨工业大学丝状真菌瑞氏木霉纤维素酶基因转录激活模式分子机制的研究刘巍峰山东大学蓝细菌光合作用捕光复合物——藻胆体中核膜连接蛋白LCM的N端结构域的张熙颖山东大学结构和功能研究重组脱硫弧菌NiFeSe氢化酶的有效翻译及其耐氧性能的分子机理研究孙俊松上海中科高等研究院宏基因组库中新双功能木聚糖酶的富集后筛选及其定向改造安迎锋沈阳农业大学新型芳香聚酮类天然产物ABX的生物合成研究瞿旭东武汉大学阿扎霉素F生物合成机制及结构多样性的比较研究孙宇辉武汉大学结核分枝杆菌DNA聚合酶的结构与功能研究王力为中国科学院生物物理研究所一种新的古菌引物合成体系的生化性质及生理功能黄 力中国科学院微生物研究所硫化叶菌染色质蛋白Cren7与Sul7d组织包装DNA的分子机制张臻峰中国科学院微生物研究所原虫模式生物布氏锥虫mRNA 3'-末端加工装置的组成及其功能的系统分析郭学敏中山大学匍枝根霉纤维素酶系组成解析及关键酶结构与功能研究汤 斌安徽工程大学激活复合体组装与顺式元件在σ70启动子上的几何分布的相关性研究王忆平北京大学水稻细菌性条斑病菌VI型分泌系统的功能鉴定何勇强广西大学构建高产紫杉醇基因工程菌株及其代谢途径与调控的研究周东坡黑龙江大学1386生命科学第24卷利用基因组发掘的方法在链霉菌SH-62中寻找新型天然产物的生物合成基因何 璟华中农业大学苏云金芽胞杆菌晶胞粘连必需基因簇scaABCDE在将伴胞晶体定位于芽胞外壁内孙 明华中农业大学侧所起的作用海洋石油细菌T7-7降解烷烃的分子机制的研究和功能优化冯 露南开大学黑曲霉ATP-柠檬酸裂解酶基因生物学功能研究刘 浩天津科技大学食线虫真菌捕食器官形成的基因组研究季星来云南大学抗菌环肽类抗生素ilamycins的生物合成研究马俊英中国科学院南海海洋研究所调节铜绿假单胞菌毒力因子表达的关键节点基因rrhS的发现及作用分子机理研究蓝乐夫中国科学院上海药物研究所hetZ基因调控丝状固氮蓝细菌细胞分化的机理徐旭东中国科学院水生生物研究所类芽孢杆菌固氮基因簇在大肠杆菌和枯草芽孢杆菌中的表达和调控研究陈三凤中国农业大学根瘤菌NGR234的III型效应因子NopL和NopM的功能性分析Christian 中山大学Staehelin微孢子虫类枯草杆菌蛋白酶的功能研究周泽扬重庆师范大学表达IBDV 主要保护性抗原的重组杆状病毒构建及其作为疫苗的免疫效果研究平文祥黑龙江大学剪切敏感型霉菌的理性选育和形态塑造及其在发酵中的应用周祥山华东理工大学常见致病军团菌O抗原基因簇分子进化机制的研究曹勃阳南开大学细菌表面多糖抗原合成基因功能的系统研究刘 斌南开大学高效合成NISIN乳酸乳球菌的简化基因组构建及其优化乔建军天津大学一种新的细菌间基因水平转移途径? ——枯草芽孢杆菌细胞间遗传重组过程的陈向东武汉大学基本特性研究及机理揭示梭菌产丁醇途径重构与调试蒋 宇中国科学院上海生命科学研究院微生物活体操纵与可控装配新方法的构建及其应用杨 光华中科技大学基于BIPES的微生物群落分析新方法研究周宏伟南方医科大学改造基因组以提高微生物生长速度的研究陈国强清华大学木糖异构酶在酿酒酵母中高效转化木糖的适配性分子机制研究鲍晓明山东大学提高由芯片合成的寡核苷酸及其组装而成的基因DNA的保真性洪 泂中国科学技术大学稳定性同位素示踪环境微生物核酸DNA的方法研究贾仲君中国科学院南京土壤研究所构建矩阵式高通量扩散室和培养方法分离培养深海沉积物中小单孢菌群郝玉有中国科学院上海生命科学研究院土壤细菌生物氧化Mn(Ⅱ)的分子机制研究李 林华中农业大学丛枝菌根真菌Glomus mosseae Gm201基因在共生早期信号转导中的生物学赵 斌华中农业大学功能研究Sphingomonas sp. DC-6降解乙草胺的代谢途径及分子机制研究何 健南京农业大学拮抗木霉菌产生敌敌畏降解酶的诱导途径与作用机制研究陈 捷上海交通大学一株弯曲假单胞菌N1的尼古丁降解机制研究唐鸿志上海交通大学城市湿地微生物群落结构与功能研究苏建强中国科学院城市环境研究所新疆火焰山热稳定、高活性细菌纤维素酶的基因克隆、定向进化及其李 刚中山大学热稳定机制解析生物电化学系统微生物生物膜形成与转录组研究邢德峰哈尔滨工业大学遗传差异及环境因子对芽孢杆菌反硝化性能与产气模式的影响胡 南南京工业大学海洋酸化对异养浮游细菌多样性及群落结构、功能的影响研究王健鑫浙江海洋学院藻对砷的生物转化及生化与分子机制研究叶 军中国科学院城市环境研究所珠江口硝化-反硝化-厌氧氨氧化耦合过程驱动的脱氮机制及关键环境洪义国中国科学院南海海洋研究所影响因素研究聚3-羟基丁酸酯(PHB)可控生物降解的研究陈 珊东北师范大学含硫废气生物过滤过程中嗜酸性微生物的发育和稳定机制李建军广东省微生物研究所苏北海岸带盐沼卤代甲烷降解菌的时空分布及其生态环境意义王睿勇南京大学基于生物电化学系统的氨氧化与反硝化偶联脱氮研究李大平中国科学院成都生物研究所硝酸盐抑制渤海湾海域油藏酸化及其微生物学机理研究:以绥中36-1 史荣久中国科学院沈阳应用生态研究所和埕北油田为例人体及家畜肠道细菌耐药基因谱及耐药基因传播途径和来源的分析朱宝利中国科学院微生物研究所水稻基腐病菌毒素zeamine的抗性基因克隆及其功能分析张炼辉华南农业大学第12期1387国家自然科学基金委员会生命科学部2012年度资助项目基于基因组学溯源技术对养殖环境携带泛耐药和毒素基因细菌气溶胶柴同杰山东农业大学的传播模式研究肠炎沙门氏菌SEF14菌毛相关功能的挖掘研究朱国强扬州大学肠出血型大肠埃希菌VI型分泌系统分泌蛋白鉴定及功能研究姚玉峰上海交通大学miR-27b/KSRP参与调控上皮细胞对病原微生物的天然免疫反应研究赵 建四川大学结核杆菌脂糖对调节性B细胞的诱导及免疫逃逸机制的研究潘 勤武汉大学胞外蛋白在铜绿假单胞菌生物被膜中的作用马旅雁中国科学院微生物研究所与活性氧相关的结核分枝杆菌异烟肼耐药机制研究米凯霞中国科学院微生物研究所CagA 抑制胃上皮细胞自噬及其在慢性胃炎发生中的作用机制研究邹全明中国人民解放军第三军医大学Retromer复合物调控稻瘟病菌致病分子机制的研究周 洁福建农林大学扁桃酸及其代谢途径对新生隐球菌减数分裂的效应与分子机制研究潘炜华中国人民解放军第二军医大学宿主不同靶器官中新生隐球菌侵袭力差异的影响机制朱红梅中国人民解放军第二军医大学水稻矮缩病毒外壳蛋白P2与水稻IAA10蛋白相互作用的研究李 毅北京大学植物Rubisco小亚基在植物病毒侵染中的作用刘玉乐清华大学大麦条纹花叶病毒(BSMV)对玉米致病性差异关键氨基酸的定位及其李大伟中国农业大学分子机制分析猪传染性胃肠炎病毒与免疫识别信号RIG-I互作机制研究任玉东东北农业大学伪狂犬病毒编码的miRNA簇在其感染过程中的表达谱及功能研究刘正飞华中农业大学家蚕细小病毒样病毒非结构蛋白NS1的表达调控及靶分子识别胡朝阳江苏大学宿主细胞DEAD-box RNA解旋酶DDX3和DDX5在猪繁殖与呼吸茅 翔南京农业大学综合征病毒感染中的作用机制研究猪札幌病毒受体鉴定研究华修国上海交通大学松毛虫T4病毒衣壳结构及病毒衣壳分子进化胡远扬武汉大学武汉野田村病毒B2蛋白抑制RNAi介导的抗病毒免疫机制的研究周 溪武汉大学负载PrP肽的树突状细胞治疗小鼠朊病毒病过程中T细胞受体库田 婵中国疾病预防控制中心病毒病预防控变化谱的研究制所杆状病毒N-WASP同源蛋白P78/83的动态调控机制及其意义王 云中国科学院武汉病毒研究所兔出血症病毒翻译起始与调控机制研究刘光清中国农业科学院上海兽医研究所B组柯萨奇病毒衣壳蛋白VP1核定位的生物学意义赵文然哈尔滨医科大学T细胞分化因子CBF-β在HIV-1 Vif形成的E3泛素化连接酶中的作用张文艳吉林大学HIV-1 Tat蛋白促进KSHV vIL-6诱导血管生成和肿瘤形成及其分子机制白志强南京医科大学热休克蛋白抑制剂抑制疱疹病毒相关恶性肿瘤的分子机制孙晓平武汉大学HBx调节CTHRC1表达的分子机制及其在介导肝癌形成中的作用研究邬开朗武汉大学SFTS 布尼亚病毒转录调控元件的鉴定及功能研究王 涛中国疾病预防控制中心病毒病预防控制所通过新的EV71抑制剂suramin靶点鉴定探索EV71病毒宿主相互作用艾德铭中国科学院上海生命科学研究院NS3蛋白N末端α螺旋结构参与HCV病毒颗粒组装的机制研究钟 劲中国科学院上海生命科学研究院风疹病毒宿主细胞受体的研究丛浩龙中国科学院微生物研究所人类细小病毒B19基因组中央polyA位点选择性使用的分子调控机制关武祥中国科学院武汉病毒研究所突变型朊蛋白致病机理李朝阳中国科学院武汉病毒研究所能克服粘膜上皮细胞极性屏障的重组痘苗病毒HIV粘膜疫苗载体的构建鄢慧民中国科学院武汉病毒研究所脑组织特异microRNA靶向EV71减毒株的构建及免疫效果评价韩剑峰中国人民解放军军事医学科学院一种新发现的登革病毒顺式作用元件DCS-PK的复制调控作用研究秦鄂德中国人民解放军军事医学科学院磷脂爬行酶1抑制HBV复制的分子机制研究杨 静中国人民解放军军事医学科学院HPV 16 E6、E7对DNA甲基化和组蛋白甲基化及乙酰化的影响及与多种蒋明军中国医学科学院蛋白酶作用机理的研究肠道病毒71型影响线粒体介导的天然免疫信号转导机制的研究雷晓波中国医学科学院病原生物学研究所MOV10蛋白抑制逆转录转座子LINE-1转座的机理研究李晓宇中国医学科学院医药生物技术研究所嗜盐古菌噬菌体SNJ1与宿主J7敌对共进化机理研究梅运军武汉工业学院大肠杆菌原噬菌体e14、rac和CPS-53中毒素-抗毒素系统的功能性研究王晓雪中国科学院南海海洋研究所噬藻体分子遗传多样性及与淡水水华蓝藻的互作研究张奇亚中国科学院水生生物研究所1388生命科学第24卷鸡毒支原体感染相关miRNAs鉴定及其分子调控机制研究彭秀丽华中农业大学衣原体感染与细胞程序性坏死机制相互作用的研究周晓辉复旦大学鹦鹉热嗜衣原体持续性感染的分子机制研究吴移谋南华大学2植物学生长素与细胞分裂素调控杨树剥皮再生过程中次生维管组织模式重建的机理研究贺新强北京大学植物类受体激酶FLS2内吞途径的研究林金星北京林业大学芦荟叶表皮结构和组分对紫外线辐射胁迫的响应李景原河南师范大学REP1基因调控水稻内稃细胞分裂分化分子调控网络的研究袁 政上海交通大学甘遂、泽漆乳汁管的发育及不同发育时期乳汁的比较蛋白质组学研究蔡 霞西北大学植物分泌体的蛋白质组与功能姜里文香港中文大学深圳研究院高直链淀粉水稻胚乳中空状和细长状淀粉粒的发育和结构特性韦存虚扬州大学木兰目花器官形态多样性与演化徐凤霞中国科学院华南植物园拟南芥REN2在花粉管极性生长过程中的功能研究傅 缨中国农业大学拟南芥珠被发育的信号网络与调控机理苟小平兰州大学调控不同生态型拟南芥离体芽再生能力差异关键基因的分离及分子机理研究李兴国山东农业大学水稻光敏色素相互作用因子OsPIFs在调控光形态建成中的作用谢先芝山东省农业科学院谷物胚乳淀粉体的发育及其与品质形成的关系顾蕴洁扬州大学SNARE蛋白复合体在花粉管顶端生长中的构成与作用李 岩中国农业大学伞形科芹亚科真芹类植物果实结构及系统学研究刘 玫哈尔滨师范大学广义报春苣苔属(苦苣苔科)系统学研究李家美河南农业大学中国榕属无花果亚属(Ficus subg. Ficus)的分类学研究李宏庆华东师范大学世界蓼属分叉蓼组(蓼科)植物的系统学研究侯元同曲阜师范大学中国葱属的分子系统学与细胞地理学研究何兴金四川大学小麦族偃麦草属的系统地位及其物种分类研究周永红四川农业大学广义假鹰爪属(番荔枝科)分类修订侯学良厦门大学爵床科马蓝属的分类修订邓云飞中国科学院华南植物园亚洲红豆属(豆科)分类学修订李世晋中国科学院华南植物园中国天南星属(天南星科)植物的分类修订胡光万中国科学院昆明植物研究所中国香茶菜属的分类修订彭 华中国科学院昆明植物研究所山茶属茶组植物的分类修订暨茶树原产地的实质性考证杨世雄中国科学院昆明植物研究所菊科紫菀属的分类学研究高天刚中国科学院植物研究所基于多学科证据探讨樟科油果樟属的系统学问题杨 永中国科学院植物研究所以系统学为基础的蔷薇科DNA条形码研究周世良中国科学院植物研究所世界槐属(豆科)的分类修订朱相云中国科学院植物研究所中国碎米蕨类(cheilanthoid ferns)的系统学研究张钢民北京林业大学中国红藻门红毛菜属(Bangia)系统分类学研究沈宗根常熟理工学院中国淡水异极藻科( Gomphonemaceae)植物的分类学研究范亚文哈尔滨师范大学东亚棉藓科植物分子系统及多样性研究王幼芳华东师范大学中国杉藻目红藻的分类、多样性及分子系统学研究王宏伟辽宁师范大学丛藓目、紫萼藓目植物抗旱特征与分子进化的研究沙 伟齐齐哈尔大学中国海洋红藻门松节藻科的分类学及其系统修订丁兰平汕头大学若尔盖湿地及其附近水域的硅藻分类生态研究王全喜上海师范大学叉蕨属(叉蕨科)的范畴及中国叉蕨属的分类学修订董仕勇中国科学院华南植物园淡水刚毛藻科的分类及系统学研究胡征宇中国科学院水生生物研究所中-日间断分布植物双花木属的种内分化与分子谱系地理研究谢国文广州大学寻求水生植物多样性格局的形成机制—以西北干旱区水生植物为例李中强湖北大学点地梅属(报春花科)的系统学与生物地理学郝 刚华南农业大学中国水生植物广布种的比较分子谱系地理学研究徐新伟武汉大学泛热带植物区系的生物地理起源与演化——以豆科羊蹄甲属为例涂铁要中国科学院华南植物园孑遗植物-莲科的残遗中心及迁移路线研究薛建华中国科学院植物研究所。

《利用重组蛋白技术提高Taq DNA聚合酶活性的研究》篇一一、引言DNA聚合酶是分子生物学领域中至关重要的酶类,其中Taq DNA聚合酶因其高效、耐热等特性,在PCR(聚合酶链式反应)技术中得到了广泛应用。
然而,天然Taq DNA聚合酶的活性往往受到多种因素的限制,如温度、pH值、酶浓度等。
为了提高其活性并拓宽应用范围,本研究利用重组蛋白技术对Taq DNA聚合酶进行改造,以期提高其活性及稳定性。
二、材料与方法1. 材料准备本研究所用的材料包括:Taq DNA聚合酶基因、表达载体、宿主菌、各种试剂及实验器材。
所有材料均经过严格的质量控制,确保实验的准确性和可靠性。
2. 方法(1)基因克隆与表达:通过PCR技术扩增Taq DNA聚合酶基因,将其克隆至表达载体中,转化至宿主菌中进行表达。
(2)重组蛋白的纯化:采用亲和层析法对表达得到的重组蛋白进行纯化,得到纯度较高的Taq DNA聚合酶。
(3)酶活性测定:通过PCR实验,测定不同温度、pH值、酶浓度下的Taq DNA聚合酶活性,分析其最佳反应条件。
(4)酶活性比较:将改造后的Taq DNA聚合酶与天然Taq DNA聚合酶进行活性比较,评估重组蛋白技术对酶活性的提高程度。
三、实验结果1. 重组蛋白的表达与纯化通过基因克隆与表达,成功获得了重组Taq DNA聚合酶。
经过亲和层析法纯化,得到纯度较高的重组蛋白。
如图1所示,SDS-PAGE电泳结果显示,重组蛋白的纯度较高,无明显杂带。
图1:SDS-PAGE电泳结果图2. 酶活性测定与比较通过PCR实验,测定不同温度、pH值、酶浓度下的Taq DNA聚合酶活性。
结果表明,在最佳反应条件下,重组Taq DNA聚合酶的活性明显高于天然Taq DNA聚合酶。
如图2所示,在95℃下反应时,重组Taq DNA聚合酶的活性提高了约30%。
同时,在pH值为8.5的条件下,其活性也得到了显著提高。
此外,在较低的酶浓度下,重组Taq DNA聚合酶也表现出较高的活性。

霉菌基因重组-概述说明以及解释1.引言1.1 概述霉菌基因重组是一种重要的生物技术手段,通过对霉菌(如酿酒酵母、曲霉等)的基因进行重新组合和调整,以获得具有特定功能的霉菌。
基因重组技术的出现为人们研究和改良霉菌开辟了新的途径。
通过基因重组,可以将某种霉菌的有益基因引入到其他霉菌中,从而使其拥有新的性状或功能。
比如,将能够合成某种有用物质的基因导入目标霉菌,使其能够高效地产生该物质。
霉菌基因重组的应用范围广泛。
在生物医药领域中,霉菌基因重组技术被广泛应用于药物的生产与改良。
通过重组技术,人们可以改造霉菌的代谢途径,提高药物产量并改善药物质量。
在农业领域中,霉菌基因重组也可以应用于生物农药的研发,通过引入抗虫基因提高农作物的抗病虫害能力。
此外,霉菌基因重组还可以用于生物燃料的生产、食品添加剂的研发等领域。
尽管霉菌基因重组技术具有广阔的应用前景,但同时也存在一些挑战和争议。
例如,基因重组可能会引发新的生物安全风险,需要进行严格的风险评估和管理。
此外,基因重组技术的操作复杂,对专业人员的要求较高,需要精确的实验设计和操作技巧。
本文将对霉菌基因重组的原理和应用进行详细介绍,以期为读者提供更多关于该领域的了解和启发。
接下来,我们将首先介绍霉菌基因重组的定义和原理,然后探讨其在不同领域的应用,最后进行总结和展望。
1.2 文章结构文章结构部分主要用于介绍本篇文章的整体组织安排,标明各个部分的主题和内容。
在本篇文章中,共有三个部分,分别是引言、正文和结论。
引言部分(1)首先概述了本篇文章的主题:霉菌基因重组。
介绍了霉菌基因重组的定义、原理和应用,并明确了本篇文章的目的。
正文部分(2)包括了两个小节。
第一小节(2.1)详细介绍了霉菌基因重组的定义和原理,解释了霉菌基因重组是指通过改变霉菌的遗传物质,将外源基因或已有基因片段重组到霉菌基因组中,从而实现某些特定功能或产生新的有用产物。
第二小节(2.2)阐述了霉菌基因重组的应用,包括生物工程领域中生产抗生素、酶和其他有用的化学物质,以及用于研究和治疗疾病等方面。
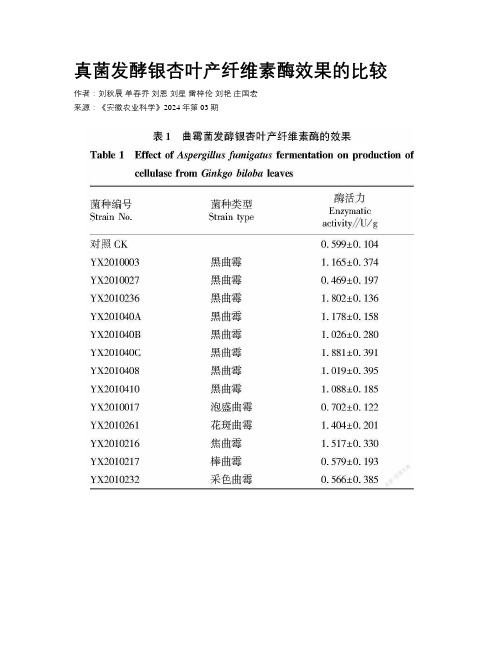
真菌发酵银杏叶产纤维素酶效果的比较作者:刘秋晨单春乔刘恩刘星雷梓伦刘艳庄国宏来源:《安徽农业科学》2024年第03期摘要为获得银杏叶饲料纤维素酶高产菌株,对从古银杏土壤中分离出的40株真菌进行发酵试验,结果表明:在供试各类真菌中,曲霉菌产生的纤维素酶分解银杏叶中的纤维素能力最强,其次是青霉,再其次是毛霉和木霉。
若用于真菌发酵产纤维素酶的生产中,曲霉和青霉是比较优良的菌种,其中编号为YX201040C的黑曲霉和编号为YX2010216的焦曲霉最为合适。
关键词真菌;纤维素酶;酶活力;葡萄糖中图分类号 S816.7 文献标识码 A 文章编号 0517-6611(2024)03-0187-03doi:10.3969/j.issn.0517-6611.2024.03.045Comparation of Cellulase Activity from Fungus Fermenting Gingko LeavesAbstract To obtain fungus that can generate cellulase productively,the fermentation experiment of 40 strains of fungi isolated from ancient ginkgo soil was carried out.According to the experiment,we found that cellulose generated by Aspergillus can degradate the cellulose in ginkgo leaf fastest,Penicillium is in the next place,mucoraceae and trichoderma are in the third place. If we want to apply fungus to generate cellulose, Aspergillus and Penicillium are good selection, among them,Aspergillus niger named YX201040C and Aspergillus ustus named YX2010216 are the best choice.Key words Fungus;Cellulase;Enzymatic activity;Glucose銀杏又称白果、公孙树,国家二级保护植物,为银杏科银杏属银杏种,属于裸子植物门,为我国独有的树种。

一种丝状真菌的荧光融合蛋白原位表达的基因方法及其应用
一种丝状真菌的荧光融合蛋白原位表达的基因方法是通过基因工程技术将荧光蛋白基因与目标基因融合,生成荧光融合蛋白基因。
然后将该荧光融合蛋白基因在真菌细胞中原位表达,以实现对目标基因在真菌细胞中的定位及可视化研究。
该方法的具体步骤包括:
1. 选择适当的真菌基因表达载体,将荧光融合蛋白基因与目标基因连接,构建融合基因表达载体。
2. 将构建好的表达载体导入真菌细胞中,通过适当的转化方法将表达载体导入真菌细胞内。
3. 利用适当的筛选标记或筛选系统,筛选出已经获得荧光融合蛋白基因在真菌细胞中的稳定表达的菌株。
4. 对所获得的荧光融合蛋白基因表达菌株进行荧光显微镜观察,并使用相关实验技术验证融合蛋白的定位及功能。
这种基因方法的应用主要是用于研究真菌细胞中特定蛋白的定位、表达及功能。
通过将荧光融合蛋白基因原位表达在真菌细胞中,可以实现对目标蛋白在细胞中的定位观察,也可以通过实时荧光显微镜观察荧光融合蛋白在细胞中的表达动态变化,从而揭示该蛋白的功能、调控以及相互作用等方面的信息。
此外,该方法也可以应用于筛选荧光融合蛋白的研究,为进一步研究和利用该蛋白提供便利。
丝状真菌瑞氏木霉生产重组蛋白的分子生物学研究进展(一)关键词:瑞氏木霉;遗传改造;基因定位置换整合;重组蛋白摘要瑞氏木霉是自然界中普遍存在并有重要经济意义的一种丝状真菌,作为工业生产菌株生产多种水解酶类已有多年历史。
本文报道了用基因工程手段对瑞氏木霉进行遗传改造,构造具新性状的重组菌株,用以过量产生同源和异源蛋白类物质的分子生物学研究进展。
包括利用CBHI 基因的强启动子在瑞氏木霉中过量表达瑞氏木霉内切葡聚糖酶、小牛凝乳蛋白酶、人抗体片段、哈茨木霉几丁质酶、Hormoconis葡萄糖淀粉酶等同源和异源蛋白以及利用在葡萄糖上强表达的启动子生产纤维素酶等遗传工程进行情况。
关键词瑞氏木霉;遗传改造;基因定位置换整合;重组蛋白1前言多年以来,丝状真菌及其产生的酶类已广泛用于食品和食品加工业。
由于其在工业上的巨大应用潜力,促进了大规模发酵工程、下游加工工程和对菌株的遗传改造的发展,其结果有助于将丝状真菌进一步应用于当今的工业生产。
大多数情况下,自然发生的菌株产生我们所需蛋白的水平很低,以至不能在商业上加以利用。
因此,为了得到所需的目的蛋白,需对自发菌株进行遗传改造。
随着分子遗传技术和真菌基因转移系统的发展,利用基因工程方法对丝状真菌进行有目的的遗传改造,已以成为可能。
丝状真菌瑞氏木霉(Trichodermareesei)是多细胞的真核微生物,其作为工业菌株用于生产分解不同植物材料的酶类,包括纤维素酶、半纤维素酶、蛋白酶、淀粉酶等,已有多年历史。
瑞氏木霉所产生的一种主要的纤维酶一纤维二糖水解酶I(cellobiohydrolase1),由单拷贝基因编码,其产量可达瑞氏木霉胞外分泌性蛋白总量的50%1]。
由此可见,cbh1启动子是很强的启动子。
因此在对瑞氏木霉的遗传改造中,常利用cbh1的启动子与终止子序列构建载体,并利用cbh1的前导肽序列引导重组蛋白进行分泌性表达。
瑞氏木霉具有极好的合成蛋白和分泌蛋白的能力;并具有真核的分泌机制,很可能还具有与哺乳动物系统相似的蛋白修饰性能,如:高甘露糖型和N-糖基化2]等。
由于瑞氏木霉具有以上优良性能,再加之其工业化规模发酵条件已比较成熟,这些都促进了对瑞氏木霉的遗传改造,为同源或异源分泌性蛋白的产生提供了一条行之有效的途径。
瑞氏木霉不仅具有适于蛋白生产的诸多优点,且对人没有毒性,在产酶条件下也不产生真菌毒素和抗生素。
近年来的实践表明,经过基因工程手术改造的瑞氏木霉重组菌株是安全无害的3]。
2过量生产同源蛋白一纤维素酶与木聚糖酶的生产纤维素酶广泛用于淀粉加工、谷物酒精发酵、麦芽制备和酿造、动物饲养以及青贮饮料加工、果汁和蔬菜汁的提取等诸多方面,近年来被应用于纺织、造纸、制浆等工业。
由于纤维素酶应用范围极其广泛,使得开发和改造纤维素酶生产菌株具有极强的现实意义。
瑞氏木霉具有降解纤维素的完全酶系,所分泌的纤维素酶混合物通常包括内切葡聚糖酶等多种酶活力。
1990年,Harkki等报道,通过基因定位置换整合使编码CBHI的基因失活,结果EGI的产量提高,不再产生CBHI4](见图1)。
1993年,Karhunen 等报道,将编码EGI的基因置于强启动子cbh1的控制之下,用此表达盒替换染色体的cbh1位点之后,EGI的产量是通常强纤维素分解菌所得CBHI量的2倍或者与其一样多5]。
其他人也曾报道过类似的情况。
瑞氏木霉的一个或几个纤维素酶基因经基因位定位置换整合而失活6,7]。
这些菌株提供了一系列令人感兴趣的新酶混合物,即可用于工业生产,又可用于研究不同酶组分对各种纤维底物的分解作用。
但是,基因失活和基因置换的方法,可能不适用于需极高特异和必须去除非必需酶活力的酶制剂。
这是因为用于酶生产的培养基,可能会诱导产生大量其它水解酶类。
而要使编码这些酶的基因都失活,在实际当中几乎是不可能的。
为了防止这类问题的产生,Goldman(1992)、Schindler(1993)Nakari(1995)等人先后报道从瑞氏木霉中分离了非纤维素酶基因的启动子,包括pgk启动子、pkiA启动子、tefl启动子和一个在葡萄糖培养基上强表达但未知的cDNA1的启动子,这些启动子可以在葡萄糖培养基上启动合成所需要的纤维素酶,产生的酶制剂基本上没有其它纤维素酶活力。
在cDNA1启动子控制下所合成的纤维二糖水解酶和内切葡聚酶产量最高可达50-100mg/L,占分泌蛋白总量50%以上8]。
1996年,Kurzatkowski等构建了能在葡萄糖培养基上生长的瑞氏木霉的重组菌株,其携带的木霉糖酶Ⅰ或木霉糖酶Ⅱ结构基因是在内源丙酮酸激酶基因(pkil)表达信号的控制之下表达。
通过SDS-聚丙烯酰胺凝胶电泳和免疫染色,发现这两种类型的转化体在葡萄糖培养基上生长时,能分别分泌出木霉糖酶Ⅰ和木霉糖酶Ⅱ。
对于木霉糖酶来说,最好的转化体产生的特异性酶活力为76U/mg;而对于木霉糖酶Ⅱ来说,则为145U/mg;都大大高出于自发菌株所产生的26U/mg9]。
3异源蛋白的生产3.1真菌酶蛋白类将TrichodermaharzianumP1染色体上的内切几丁质酶基因分离,导入丝状真菌瑞氏木霉,并在cbh1启动子过量表达。
出发菌株瑞氏木霉RutC-30不具有任何内切几丁质酶活力。
T·harzianum内切几丁质酶基因编码区前的区段的瑞氏木霉中能正确发挥功能。
摇瓶培养产生130mg/L 有活力的几丁质酶,比T·harzianum产生的内切几丁质酶活力提高大约20倍。
瑞氏木霉RutC-30似乎能忍受内切几丁质酶具有抗植物病原真菌的活力10],已被用于植物病源真菌的生物防治。
另据B.Cubero等(1997)报道,在瑞氏木霉组成型启动子ki和cbh2终止子的控制之下构建了两种载体,分别含有T·harzianum的基因chit33和bgn16.2的开放阅读框(chit33编码一种内切几丁质酶,bgn16.2编码一种外切葡萄糖淀粉酶),并获得了稳定的拷贝转化体。
对转化体bng16.2来说,整合到基因组中的基因拷贝数与mRNA和蛋白质的积累量和在葡萄糖阻遏或几丁质酶诱导条件下的胞外牧特性酶活力之间均呈正相关。
该菌株比野生型菌株能更迅速地抑制病原真菌的生长。
对转化体chit33来说,在葡萄糖阻遏的条件下。
对转化体chit33来说,在葡萄糖阻遏的条件下,其产生的胞外几丁质酶活力是野生菌株的10倍11]。
JoutsjokiVV(1994)报道,构建了两种一步基因置换载体。
一种载体含有Hormoconisresinae葡萄糖淀粉酶P的基因组基因(gamP),另一种含有相应的cDNA,都在瑞氏木霉cbh1启动子控制下表达。
这些载体经转化后置换瑞氏木霉的cbh1基因。
在这两种载体中,cbh1启动子都能精确地连接到gamP蛋白质编码区上游,指导瑞氏木霉分泌出有活力的葡萄糖淀粉酶P(GAMP)。
这表明gamP基因中内含子序列在瑞氏木霉中得到了正确的加工。
研究结果表明,含有gamPcDNA的最优转化体能分泌大约700mg/L有活力的GAMP,是在H.resinae中产量的20倍12],但chb1基因被gamP基因组基因置换的瑞氏木霉转化体,所分泌的有活性的GAMP要多于含有3拷贝cDNA的转化体。
对瑞氏木霉分泌H.nisresinae葡萄糖淀粉酶P的研究结果表明,自然状态未成熟GAMP 的N端延伸部分可以在瑞氏木霉中作为一种有效的分泌信号,所产生的胞外葡萄糖淀粉酶活力高于以CBHI信号肽作为分泌信号时的情况13]。
酵母基因在瑞氏木霉中表达的一例是:酿酒酵母(Saccharomycescerevisiae)DPM1基因(编码甘露糖基磷酸多萜醇合成酶)的编码区插入到pLMRS3质粒中,在瑞氏木霉pki启动子和cbh2终止子控制下表达。
在pFG1质粒(含有瑞氏木霉pyr4基因)存在下,与pLMRS3质粒一起对瑞氏木霉的一种pyr4阴性突变株TU-6进行共转化。
然后筛选pyr4阳性转化子,得到4株多拷贝DPM1基因串联整合的稳定转化子。
与受体菌株TU-6相比,转化子的甘露糖基磷酸多萜醇合成酶活力提高了15-19倍,分泌的蛋白总量也提高了4倍14]。
除以上蛋白外,被表达的真菌蛋白还包括肌醇六磷酸酶等。
这些酶都是在cbh1启动子下表达,摇瓶培养产量达100mg/L水平。
发酵培养中,产量为每升几克。
在cbh1启动子的控制之下,来自担子菌纲的真菌Phlebiaradiate的氧化还原酶(如:漆酶)的产量也达到中等水平3.2哺乳动物蛋白瑞氏木霉已被用于牛凝乳蛋白酶的生产。
利用cbh1启动子和终止子,通过共转化外源基因与来自构巢曲霉(Asp.nidulans)的amdS基因一起引入乙酰胺非利用型菌株。
测量到的凝乳酶mRNA和分泌出的凝乳酶的量要比相应的cbh1mRNA和CBHI的量低1-2个数量级。
这表明牛凝乳蛋白酶在瑞氏木霉中的表达受到了转录限制。
但是,所获得的40mg/L的水平,与典型的在构巢曲霉素中的2-10mg/L的最初水平相比,还是一个可喜的进步15,16]。
另外,据文献报道,利用其它丝状真菌生产的几种人体蛋白是:表皮生长因子(hEGF),生产激素(hGH),白细胞介素6,组织血纤维蛋白溶酶原激活因子(htPA)等。
Fab分子是由抗体完整的轻链和重链的Fd部分,由链间二硫链连接在一起。
这种分子曾由木霉成功地生产出来,并用于研究其分泌过程。
利用cbh1启动子和信号序列,分别构建了表达轻链和重链的表达盒。
首先构建的是只产生轻链的菌株,然后用2种不同的重链表达载体进行再转化。
这两种载体分别指导Fd链或者与CBHI核心-连接区融合在Fd链的表达。
只产生轻链的菌株分泌的轻链水平很低(0.2mg/L)。
对前一种重链载体的转化体来说,在摇瓶培养中,分泌的有可能的Fab 分子为1mg/L。
在具有cbh1-Fd融合结构的转化体菌株中,观察到产量有显著增长。
最优菌株能分泌40mg/L具有免疫活力的CHBI-Fab融合蛋白。
在发酵培养中,水平增长到150mg/L.然而,没有融合到CBHI上的Fab分子,水平仍为1mg/L。
有趣的是,一种未知的木霉蛋白酶在CBHI-Fab融合蛋白的N-端特异去除2个氨基酸,能产生少量的Fab分子。
释放的Fab分子表现出免疫活性和对抗原的亲和性,而CBHI-Fab 分子的免疫活性要低5-12倍。
对分离到的抗体链进行定量测定表明,分泌出的所有轻链都和重链组装在一起。
CBHI-Fab产物的上清液中,含有显著数量(800mg/L)的CBHI核心-连接区肽,它们来自CBHI-Fd 融合分子。
这样,CBHI-Fd的产生是非常有效的(大于800mg/L),但其中只有少量(40mg/L)装配成有功能的CBHI-Fab分子(或者,释放的Fab分子)。