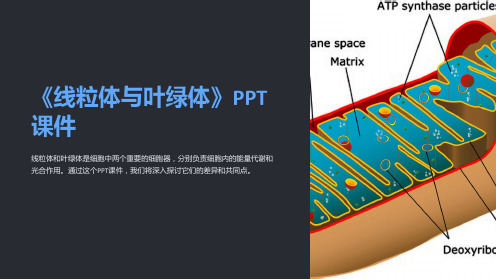
线粒体和叶绿体是半自主性细胞器,有自己的环状 DNA和转录、翻译体系,是真核细胞的第二遗传信 息系统。
ppt课件
2
第一节 线粒体与氧化磷酸化
线粒体是细胞的动力工厂:
•人体细胞内95%的ATP在线粒体产生; •线粒体通过氧化磷酸化进行能量转换; •为细胞各种生命活动提供能量。
ppt课件
3
ppt课件
2. 衰老:随年龄的增长,细胞内线粒体数量减少而体积增大, 损伤的mtDNA的积累越来越多。线粒体产生的氧自由基损伤 DNA,而线粒体中无DNA损伤的修复系统。由mtDNA 编码 的酶的结构改变,功能异常。
3. 细胞凋亡:凋亡信号引起线粒体释放细胞色素c, 参与凋亡 信号的传导。
ppt课件
28
第二节 叶绿体与光合作用
ppt课件
11
二、线粒体的功能
• 人体细胞95%的ATP来源于线粒体。是物 质彻底氧化分解的场所。
还参与氧自由基的生成;细胞凋亡的调控;细 胞中Ca2+的稳态调节等。
ppt课件
12
线粒体中的氧化代谢
蛋白质,糖类和脂肪酸最终分解为乙酰CoA,进入 TCA(tricarboxylic acid cycle) 循环,产生NADH或 FADH2。
(一)叶绿体膜
• 双层膜组成,膜间为10~20nm的间隙。
• 外膜的通透性大,内膜通透性低;ADP、
ATP、 NADP+、葡萄糖和焦磷酸等需要特
殊的转运体才能通过内膜。
NADP+,烟酰胺腺嘌呤二核苷酸磷酸,是氧化型。NADPH是还原型 (辅酶II)
ppt课件
31
(二) 类囊体
• 是由内膜发展而来的封闭的扁平囊。 • 类囊体膜:流动性高,促进光和作用的酶类复合