
细胞周期的关键分子调节机制
摘要:细胞周期是指连续分裂的细胞从一次有丝分裂结束到下一次有丝分裂完成所经历的整个序贯过程. 细胞周期中每一事件都是有规律、精确地发生, 并且在时间与空间上受到严格调控. 细胞周期中最关键的三类调控因子是: cdc 基因、周期蛋白依赖性激酶( CDKs) 及细胞周期蛋白( cyclin) . 这些调控因子的发现对肿瘤学及发育生物学的发展都有重要的理论和实践意义.
关键词:细胞周期, cdc 基因, 周期蛋白依赖性激酶, 细胞周期蛋白
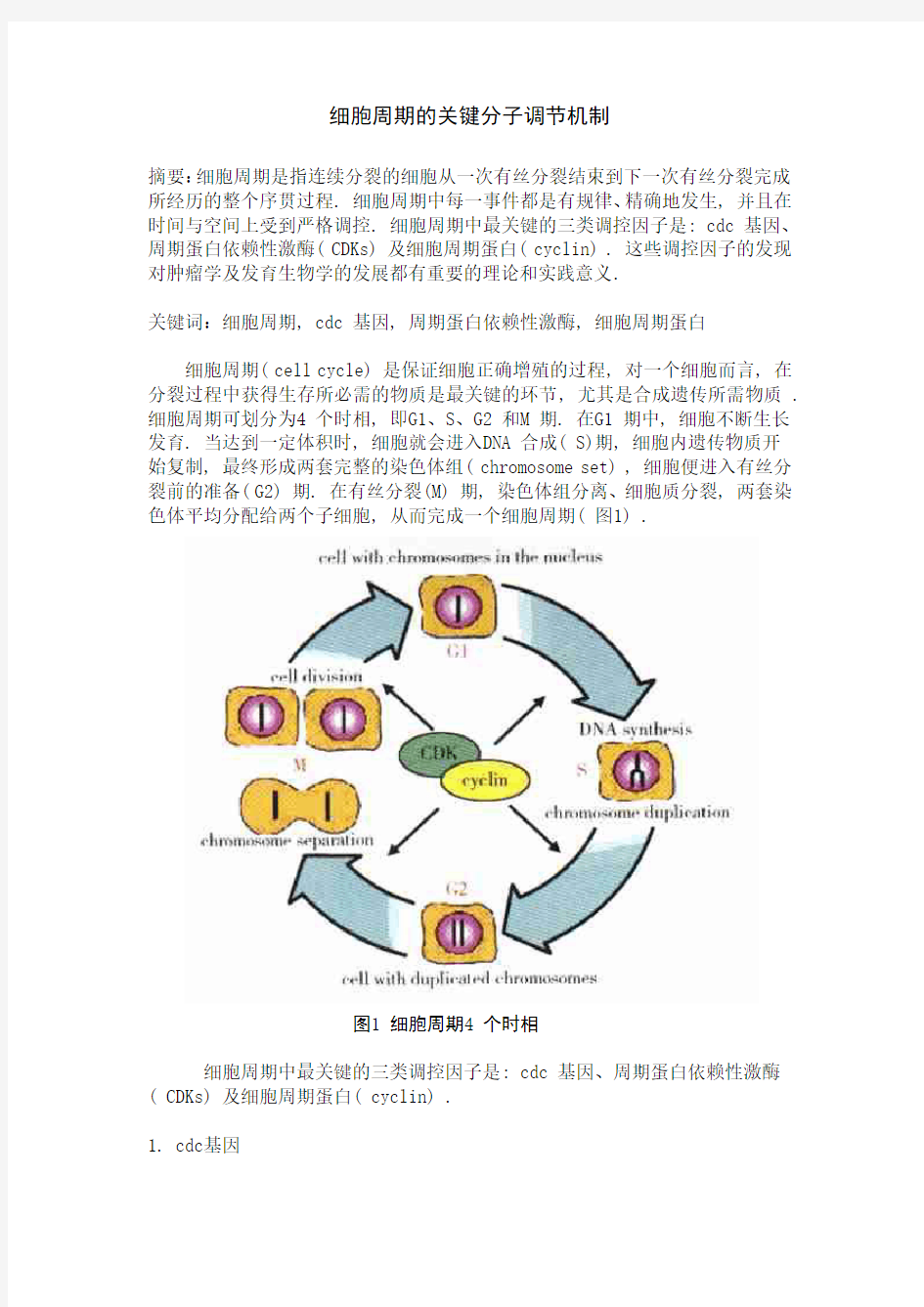
细胞周期( cell cycle) 是保证细胞正确增殖的过程, 对一个细胞而言, 在分裂过程中获得生存所必需的物质是最关键的环节, 尤其是合成遗传所需物质 . 细胞周期可划分为4 个时相, 即G1、S、G2 和M 期. 在G1 期中, 细胞不断生长发育. 当达到一定体积时, 细胞就会进入DNA 合成( S)期, 细胞内遗传物质开
始复制, 最终形成两套完整的染色体组( chromosome set) , 细胞便进入有丝分裂前的准备( G2) 期. 在有丝分裂(M) 期, 染色体组分离、细胞质分裂, 两套染色体平均分配给两个子细胞, 从而完成一个细胞周期( 图1) .
图1 细胞周期4 个时相
细胞周期中最关键的三类调控因子是: cdc 基因、周期蛋白依赖性激酶( CDKs) 及细胞周期蛋白( cyclin) .
1.cdc基因
1.1 cdc基因的发现
哈特韦尔采用遗传学方法, 用芽殖酵母( Saccha rymyces cerevisiae)作为实验对象研究细胞周期.20世纪70年代初,他通过温度敏感突变技术筛选出突变酵母细胞, 这些细胞的生长停滞在特定的细胞周期时相 , 从而确定缺陷基因所编码的蛋白质在细胞周期调控中的作用,利用种方法,他成功地分离出上百个涉及细胞周期调控的基因(图2),并命名为cdc 基因.
图2 用荧光钙( calcofluor) 示裂殖酵母(Schizosacchharomyces
pombe) 细胞壁和中隔( septum)
野生型细胞的长度加倍并一分为二, 而cdc25 缺陷的细胞已长的很长却不分裂. cdc25 是细胞从G2 期进入M 期必需的基因, 它负责CDK2 的去磷酸化( 引自Nurse P) .
1.2 cdc基因的功能
在哈特韦尔发现的这类基因中, cdc4、6、7、8 等控制DNA 复制, 如cdc8 具有起始DNA 合成的功能 ; cdc5、14、15 等参与染色体分离的调控; cdc3、10、11、13 等调控细胞质的分裂,名为cdc28 的基因, 启动细胞从G1 期进入S期. 该基因编码的蛋白质是其他cdc 基因产物执行功能的前提, 所以又被称为star t基因。
2.周期蛋白依赖性激酶(CDK)
2.1 CDK的发现
20 世纪70 年代中期, 纳斯采用遗传学方法和分子生物学方法, 在对裂殖酵母
的细胞周期进行研究时, 从中发现了cdc2 基因, 该基因在细胞分裂调控中起着关键作用, 它控制细胞从G2 期进入M 期.罗纳斯又发现cdc2 基因还有着与
start 基因相同的作用,即在细胞从G1 期向S 期的转换进程中起调控作用。1987 年, 保罗纳斯从人体中发现了细胞周期的一个关键调节物质CDK( 周期蛋白依
赖性激酶cyclindependent kinase) , 并发现CDK 在控制细胞分裂( 从G2期到有丝分裂期) 中有关键作用, 它可使其他蛋白质磷酸化, 从而使细胞沿着细胞周
期各时相发育, 不断地分裂增殖。CDK 在进化中高度保守.
2.2 CDK的功能
CDK的主要生物学作用是启动DNA的复制和诱发细胞的有丝分裂, 从而驭动细胞周期, 在细胞发育过程中起着非常重要的作用。研究表明:这种激酶的周期性激活与失活是推动细胞周期进行的主要因素。
2.3 CDK激酶的激活
CDK激酶的激活作用依赖于可逆的蛋白质磷酸化作用, 即通过磷酸基与靶蛋白结合来构筑这种生命体代谢过程中最基本的公共通路。靶蛋白的磷酸化改变了靶蛋白的功能, 使它处于高度活跃或完全静止的状态。由于激酶把碑酸醋涂抹在许多不同的靶蛋白上, 从而可同时改变细胞中的不同进程, 将高强度的细胞分裂信
号传遍细胞内部。
3.周期蛋白(cyclin)
3.1 周期蛋白(cyclin)的发现
20 世纪80 年代早期,蒂莫西亨特采用国王海胆( A rbacia) 作为实验模型, 第一个发现了周期蛋白( cyclin) 分子。由于其表达水平随细胞周期而剧烈起伏, 而被命名为周期蛋白.该蛋白在细胞周期中周期性地生成, 然后降解。
3.2 周期蛋白(cyclin)的功能
周期蛋白(cyclin)与CDK 分子结合, 从而对CDK 的活性进行调控, 选择性地将磷酸基团连接在某些蛋白质分子上。周期蛋白的周期性降解, 是细胞周期调控的一个重要机制。不同的cyclin分别在某一个细胞周期时相表达, 又在另一个时相降解 , 这种性质是细胞周期调控的一种独特现象. cyclin 这种独特的性
质被亨特察觉, 通过进一步研究, 他发现细胞周期的运转必需有cyclin 的参
与 , cyclin 缺陷型细胞停滞在G1 或G2 期, 无法进行有丝分裂.
4. 细胞周期调控的分子机制
在多细胞真核生物中,参与细胞周期调控的核心蛋白分子主要分为3大类,分别是:周期蛋白依赖性激酶(CDK)、周期蛋白(Cyclin)及“细胞周期蛋白依赖性激酶抑制因子”(Cdk inhibitor,CKI)。其中,Cdk是细胞周期调节的中心环节,Cyclin是Cdk的正调节因子,CKI是Cdk的抑制因子。
4.1细胞周期的驱动和调控机制
作为“细胞周期发动机”,Cdk在细胞周期的调节中起关键作用。各种Cdk在细胞周期的各个特定时问被激活,通过磷酸化底物,驱使细胞完成细胞周期。这就是细胞周期的驱动机制。(图 3)
图3 S期起始调控的分子机制示意图
4.2 在整个细胞周期过程中,细胞内各种Cdk的含,即活化的Cdk与非活化的Cdk 的总量不变,改变的只是它们之间的比例。这个比例的改变主要受三方面的调节:(1)Cdk只有通过与特定的Cyclin形成二聚复合物才能发挥作用
(2)Cdk的激活还需要在其保守的苏氨酸和酪氨酸残基上发生磷酸化
(3)Cdk的活性可被Cdk抑制蛋白(CKI)抑制
4.2.1 Cdk只有通过与特定的Cyclin形成二聚复合物才能发挥作用
目前,在高等真核细胞中Cyclin主要包括8个成员(A~H),它们在细胞周期的不同时相,结合不同的Cdk发挥不同的作用。根据Cyclin发挥作用时相的不同,通常将其分为四类:G一Cyclin、G/S—Cyclin、S-Cyclin和M—Cyclin.
图4 各种cyclin-cdk复合物作用的细胞周期时相
在结构上,所有的Cyclin分子均有一个相对保守区域,称为细胞周期蛋白盒(Cyclin box),其主要功能是与Cdk结合而改变Cdk的蛋白质构象,激活Cdk的蛋白激酶活性,具有激酶活性的Cdk使周期蛋白特定的氨基酸残基磷酸化,使后者的三维构象发生变化,从而引起一系列的链式反应,调控细胞周期进程.
4.2.2 Cdk的激活还需要在其保守的苏氨酸和酪氨酸残基上发生磷酸化
这个磷酸化是由Cdk激活激酶(Cdk—activa—ting kinase,CAK,即Cdk7/Cyclin H)完成的。CAK可以磷酸化Cdkl的Thr一161位点,使其活化,改变Cdk的分子构象,促进Cdk与Cyclin结合,所以促进了细胞周期的进程.
4.2.3 Cdk的活性可被Cdk抑制蛋白(CKI)抑制
CKI通过直接结合Cdk,或与Cdk—Cyclin复合体作用,抑制Cdk的作用,调节细胞周期。
目前,将CK1分为两大家族
⒈具有广泛抑制Cdk作用的“Cdk抑制蛋白/激酶抑制蛋白”家族
⒉具有特异性抑制作用的“Cdk 4抑制因子”
因此,正常的细胞周期需要Cdk的正调节因子Cyclin与负调节因子CKI的精确协同与平衡,一旦这种平衡失稳就会造成细胞的失控性增殖,发生癌变。
4.3 细胞周期的监控机制——“检验点”机制
细胞周期是高度有组织和精确的时序调控过程,它严格地沿着G 一S—G 一M 的顺序循环运转,为保证这一过程的正常进行,细胞形成一套检验细胞周期中DNA 合成和染色体分配的机制,即“细胞周期检测点”。这些监测机制可以检测到DNA 结构的受损或复制不全,还能检测到细胞分裂过程中所需的蛋白复合物的缺失。在酵母细胞中,当发生影响细胞周期正常运行的事件时,检测点相关的信号转导通路将被激活,它们可以使Cdc 2p酪氨酸残基持续的磷酸化而抑制Cdk的活化,从而阻止有丝分裂期的发生;或通过其它机制在有丝分裂期的较晚时期阻断细胞
周期的继续进行。在哺乳动物细胞中,当DNA损伤时,还可通过p53基因的作用将细胞周期阻断于s期。阻断细胞周期的进行后,“检测点”机制采取诱导基因转录、促进DNA的修复等有效的补救措施以排除故障。当故障排除后,细胞周期才能恢复运转。当损伤过大,细胞无法修复时,检测点将启动细胞凋亡程序以清除那些带有病变倾向的细胞,减少对机体的危害。
目前为止可以将所发现的细胞周期检测点分为三种:
⒈DNA损伤检测点,它包括两个关键性检测点:G /S转换点和G /M转换点
⒉DNA复制检测点,其在s期中负责DNA复制的进度
⒊纺锤体组装检测点,其在分裂期起作用,检测纺锤体有无组装、染色体是否正确排列并与纺锤体连接,以及染色体是否正确分配等
这些检测机制保证了DNA在分子水平上的精确复制及在细胞水平上的精确分离检测点对细胞周期进程进行严格的监督,使DNA复制和有丝分裂准确无误地进行,保证遗传的稳定性。它们的缺失将导致细胞在没有正确完成前一时相就进入下一时相,细胞将出现严重的遗传性损伤甚至癌变,最终导致机体死亡。
5.现状前景及仍存在的问题
5.1 现状前景
细胞周期调控的分子机制的揭示,将有助于人们理解肿瘤细胞中染色体是如何进行重排、丢失或不均等地分配到子代细胞中, 从而使人们有可能更好地对肿瘤进行预防、诊断及治疗。
5.2 仍存在的问题
细胞增殖, 尤其是它的调控机制仍远未彻底阐明,譬如:参与周期调控的基因绝非仅此几个, 它们在促进或是阻止周期进行中究竟起何作用, 它们与START、CDK 有何关系, 高等动物细胞与酵母的周期调控有何差别等问题, 都值得人们
进一步思索与探讨。
参考文献
1 Nurse P. Regulat ion of the eukaryot ic cell cycle. European JCancer, 1997, 33 ( 7) : 1002~ 1004
2 Bart let t R, Nurse P. Yeast as a model syst em for un derst anding the cont rol of DNA replication in eukaryot es. Bioessays, 1990,12 ( 10) : 457~ 463
3 Hartwell L H. Macromolecule synthesis in temperature2sensit ivemut ant s of yeast . J Bact eriology, 1967, 93 ( 5) : 1662~ 1670
4 Hartwell L H. Cell division from a genetic perspect ive. J Cell Biol,1978, 77: 627~ 637
5 Hartwell L H. Genet ic cont rol of the cell division cycle in yeast ò:genes cont rolling DNA replicat ion an d it s init iat ion. J Mol Biol,
1971, 59: 183~ 194
6 Reid B J, Hartwell L H. Regulation of mat ing in the cell cycle of Saccharymyces cerevi si ae. J Cell Biol, 1977, 75: 355~ 365
7 Evans T , Rosenthal E T, Hunt T , et al . Cyclin: aprot ein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division. Cell, 1983, 33: 389~ 396
8 Hunt T , Luca F C, Ruderman J V. T he requirements for prot ein synthesis and degradat ion, and the cont iol of dest ruct ion of cyclins A and B in the meiot ic an d mitotic cell cycles of the clam embryo. J Cell Biol, 1992, 116 ( 3) : 707~ 724
9 Hartwell L H, Culott i J, Pringle J R, et al . Genetic cont rol of the cell division cycle in yeast. Science, 1974, 183: 46~ 51
10 Hunt T . Cyclins an d th eir partners: from a simple idea to complicat ed reality. Semin Cell Biol, 1991, 2 ( 4) : 213~ 222
11 Ban dara L R, Adamczewski J P, Hunt T , et al . Cyclin Aand the retinoblastoma gene product complex with a common t ransct ipt ion factor. Nature, 1991, 352 ( 6332) : 249~ 251
12 Nurse P, T huriaux P. Regulatory genes cont rolling mitosis in the fission yeast Schizosacchharomyces pombe. Genet ics, 1980, 96( 3) : 627~ 637
13 Simanis V, Hayles J, Nurse P. Control over the onset of DNA synthesis in fission yeast. Philos Trans R Soc Lond B Biol Sci,1987, 317 ( 1187) : 507~ 516
14 Nurse P. Universal cont rol mechanism regulat ing onset of M2 phase. Nature, 1990, 344 ( 6266) : 503~ 508
15 Simanis V, Nurse P. T he cell cycle control gene cd c2 + of fission yeast en codes a prot ein kinase pot ent ially regulated by phosphorylat ion. Cell, 1986, 45 ( 2) : 261~ 268
16 Li A, Blow J J. T he origin of CDK regulat ion. Nature Cell Biology, 2001, 3: 182~ 184
17 Gould K L, Nurse P. T yrosine phosphorylat ion of the fission yeast cdc2+ protein kinase regulat es ent ry into mitosis. Nature, 1989, 342 ( 6245) : 14~ 15
18 Fel ix M A, Labbe J C, Dorse M, et al . T riggering of cyclin degradation in int erphase extracts of amphibian eggs by cdc2 kinase. Nature, 1990, 346 ( 6282) : 379~ 382
19 Hartwell L H, T ed A. Weinert Chekpoints: Controls that ensure the order of cell cycle events. Science, 1989, 246: 629~ 633
20 Paulovich A G, Hartwell L H . A checkpoint regulat es the rat e of progression through S phase to DNA damage. Cell, 1995, 82: 841~ 847 21 Paulovich A G, Armour C D, Hartwell L H. The saccharomyces cerevisiae RAD9, RAD17, RAD24 and MEC3 genes are required
for tolerat ing irreparable, ult raviolet2induced DNA damage. Genet ics, 1998, 150 ( 1) : 75~ 93
22 DpUrso G, Grallert B, Nurse P. DNA polymerase alpha, a component of the replicat ion initiat ion complex, is essential for the checkpoint coupling S to mitosis in fission yeast . J Cell Sci, 1995,108: 3109~ 3118 23 Hartwell L H, Szankasi P, Robert C J, et al . Integrating genet ic approach es into the discovery of ant icancer drugs. Science, 1997,278 ( 5340) : 1064~ 1068
24 Zhang P. The cell cycle and development : redundant roles of cycle regulators. Current Opion in Cell Biology, 1999, 11: 655~ 662
25 齐云. 发现调控细胞循环周期的神秘物质. 世界科学,2001, 11: 7—8.
26 颜亮, 陈勇. 生理学或医学奖: 加深对生命基本过程的理解. 大众科技报, 2001—10—14( 5) .
27 刘元亮, 姚慧华, 冠世琪等. 科学认识论和方法论. 北京:清华大学出版社, 1987.
28 陈衡. 科学研究的方法论. 北京: 科学出版社, 1982.
29 郝水. 细胞如何调控基因组的有序活动. 李喜先. 21 世纪100 个科学难题. 长春: 吉林人民出版社, 1999, 607—614.
30 蔡如鹏. 细胞“引擎”驱动癌症研究. 科学时报, 2001,( 6) : 10—16.
31 申景平, 卢圣栋. 癌症的靶向性基因治疗. 世界科学,2000, 1: 31.
32 Dan Ferb er, 李伟. 与癌症治疗副作用抗衡的新疗法. 世界科学, 2000, 1:
33 刘擎, 余龙. 酵母: 一种模式生物. 生命的化学, 2000;20(2) : 61
34 赵炬才, 张广泉. 细胞周期调控与骨肉瘤. 中华骨科杂志, 2000;20: 442 46
35 吴家睿. 检查点: 细胞周期的质量监督. 生命的化学, 1999; 5: 1992202
36 SerranoM. ,Hannon G. J. , Beach D. , et al . A new regulatory mot if in cell cycle control causing specific inhibition of cyclin DPCDK4. Nature,1993; 366( 16) : 704
37 KondoT. , et al . Mol . Cell . Biol . , 1999;19: 1136
38 Shimomura T. , et al . Mol . Cell . Biol . , 1998; 18: 5485
39 Navas T. A. , et al . Genes Dev. , 1996;10:2632
40 Sugimoto K. , et al . Mol . Cell . Biol . , 1997; 17: 5905
41 Hartwell L. H. , Kastan M. B. Cell cycle control and cancer. Science,1994; 266: 182121828
42 Nobori T. ,Miura K. , Wu D. J. , et al . Delet ions of the cycl in dependent kinase 4 inhibitor gene in mult iple human cancers. Nature, 1993; 368:753
43 Hunt er T. , Pines J.Cycl ins and cancer ò, cyclin D and CDK inhibitors come of age. Cell , 1994; 79: 5732582
44 赵晓荣, 顾焕华, 翁新宪等. 细胞周期蛋白( cyclin) D 在鼻咽癌中的表达及功能初步分析. 生物化学与生物物理学报, 2000; 32( 2) :1922196
45 OhtsuboM. , Thoedoras A.M. ,Schumacher J. , et al .Human cyclin E. ,
a nuclear protein essent ial for the G12to2S phase transition. Mol . Cell . Biol . , 1995; 15:261222624
46 Girard F. ,Strausfeld U. , Fernandez A. , et al . Cyclin A is required for the onset of DNA repl ication in mammalian fibroblasts. Cell , 1991; 67:116921179
47 Harris H.The Birth of the Cel1.New Haven:Yale Universi—ty Press,1999.
48 Nurse P.A long twentieth century of the cell cycle and be— yond.Cell,2000,100 :71—78
49 Howard A ,Pelc S. Synthesis of deoxyribonucleic acid in normal and irradiated cells and its relation to chromosome breakage. Heredity,1953,6 :261—273.
50 Hartwell LH,Mortimer RK,Culotti J,et a1.Genetic control of the cell division cycle in yeast:genetic analysis of cdc mutants.Genetics,1973,74:267—286.
51 L.Hartwel1.Saccharomyces cerevisiae cell cycle. Bacteriol Rev,1974,38 :164 ~198.
52 Draetta G,Beach D.Activation of cdc2 protein kinase dur— ing mitosis in human cells:cell cycle-dependent phosphoryl— ation and subunit rearrangement.Cell,1988,54 :17—26.
53 Simanis V.Nurse P. The cell cycle control gene cdc2 of fission yeast encodes a protein kinase potentially regulated by phosphorylation.Cell,1986,45 :261~268.
54 Nurse P.Cell cycle control genes in yeast.Trends Genet.1985.1:51—55.
55 Russell P,Nurse P. Negative regulation of mitosis by weel ,a gene encoding a protein kinase homolog.Cell,1987,49 :559~567.
56 Russell P,Nurse P.cdc25 functions as an inducer in the mitotic control of fission yeast.Cell,1986,45 :145 ~153.
57 Russell P,Nurse P. The mitotic inducer niml functions in a regulatory network of protein kinase homologs control—ling the initiation of mitosis.Cell,1986,49 :569~576.
58 NUrse P.Universal control mechanism regu lating onset of M phase.Nature,1990,344 :503~508.
59 Lee MG,Nurse P.Complementation used to clone a human homologue of the fission yeast cell cycle control gene cdc2.Nature,1987,327 :31~35.
60 van den Heuvel S,Harlow E.Distinct roles for cyclin-de— pendent kinases in cell cycle contro1. Science,1993,262 :2050~2054.
61 Evans T,Rosenthal ET,Youngblom J,et a1. Cyclin:a protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division.Cell,1983,33 : 389 396.
62 Hulleman E,Boonsha J.Regulation of Gl phase progres—sion by growth factors and the extracellular matrix.Cell Mol
Life Sci.2001.58 :80~93.
63 Kadis P.The cdk—activating kinase(CAK):from yeast to mammals.Cell Mol Life Sci.1999.55 :184 —296.
64 Hagopian JC,Kialey MP,Stevenson LM,et a1.Kinetic basis for activation of cdk/cyclin A by phosphorylation.Bi—ol Chem ,2001,276 :
275~280.
65 Parker LL,Piwnica—W orms H.Inactivation of the p34cdk2一cyclin B complex by the human W EE1 tyrosine kinasis.Science,1992,257 :1955~1957.
66 Abfieu A ,Brassac T,Galas S,et a1.The polo—like kinasis Pixl is
a component of the MPF amplification loop at the G2/ M—phase transition of the cell cycle in Xenopus eggs. Cell Sci,1998.111 :1751~1757.67 Chen J,Willinghan T,Shford M,et a1.Tumor suppression and inhibition if aneuploid cell accumulation in human brain tumor cells by actopic over expression if the cyclin— dependent kinasis inhibitor 27kip1. J Clin Invest,1996,97 :1983~1988.
68 Serrano M,Hannon GJ,Beach D.A new regulatory motif in cell—cycle control causing specific of cyclinD/CDK4.Na—ture,1993,366 :704 —707.
69 Hartwell L,W einert T.Checkpoints:controls that ensure the order of cell cycle events.Science。1989.246:629—634.
70 Nakayama KI,Hatakeyama S,Nakayama K.Regu lation by proteolysis of cyclin E and p27kip1. Biochem Biophys Res Commun,2001,282 :853~860.71 Xie S,Xie B, Lee MY ,et a1. Regu lation of cell cycle checkpoints by polo—like kinases.Oncogene,2005,24 :277—286.
72 Ducat D,Zheng Y.Aurora kinases in spindle assembly and chromosome segregation. Experimental Cell Research,2004.301:6O~67.
73 Foster I. Cancer:a cell cycle defect.Radiography ,2008,
结合着下载的资料复习吧~~~~ 绪论 分子生物学的发展简史 Schleiden和Schwann提出“细胞学说” 孟德尔提出了“遗传因子”的概念、分离定律、独立分配规律 Miescher首次从莱茵河鲑鱼精子中分离出DNA Morgan基因存在于染色体上、连锁遗传规律 Avery证明基因就是DNA分子,提出DNA是遗传信息的载体 McClintock首次提出转座子或跳跃基因概念 Watson和Crick提出DNA双螺旋模型 Crick提出了“中心法则” Meselson与Stah用N重同位素证明了DNA复制是一种半保留复制 Jacob和Monod提出了著名的乳糖操纵子模型 Arber首次发现DNA限制性内切酶的存在 Temin和Baltimore发现在病毒中存在以RNA为模板,逆转录成DNA的逆转录酶 哪几种经典实验证明了DNA是遗传物质? (Avery等进行的肺炎双球菌转化实验、Hershey 利用放射性同位素35S和32P分别标记T2噬菌体的蛋白质外壳和DNA) 第二章染色体与DNA 第一节染色体 一、真核细胞染色体的组成 DNA:组蛋白:非组蛋白:RNA = 1:1:(1-1.5):0.05 (一)蛋白质(组蛋白、非组蛋白) (1)组蛋白:H1、H2A、H2B、H3、H4 功能:①核小体组蛋白(H2A、H2B、H3、H4)作用是将DNA分子盘绕成核小体
②不参加核小体组建的组蛋白H1,在构成核小体时起连接作用 (2)非组蛋白:包括以DNA为底物的酶、作用于组蛋白的酶、RNA聚合酶等。常见的有(HMG蛋白、DNA结合蛋白) 二、染色质 染色体:分裂期由染色质聚缩形成。 染色质:线性复合结构,间期遗传物质存在形式。 常染色质(着色浅) 具间期染色质形态特征和着色特征染色质 异染色质(着色深) 结构性异染色质兼性异染色质 (在整个细胞周期内都处于凝集状态)(特定时期处于凝集状态)三、核小体 由H2A、H2B、H3、H4各2 分子组成的八聚体和绕在八聚体外的DNA、一分 子H1组成。八聚体在中央,DNA分子盘绕在外,由此形成核心颗粒。,H1结合在核心颗粒外侧DNA双链的进出口端,如搭扣将绕在八聚体外DNA链固定,核心颗粒之间的连接部分为连接DNA。 核小体的定位对转录有促进作用
2001年诺贝尔生理学和医学奖
细胞周期调控 一、背景介绍 2001年诺贝尔生理学医学奖授予美国西雅图弗瑞德·哈钦森癌症研究中心的Leland H Hartwell、英国伦敦皇家癌症研究基金会的Sir Paul M. Nurse和R. Timothy Hunt,以表彰获奖者们在细胞周期调控方面的卓越发现和贡献。 Leland (1939年生)在上世纪60年代末便认识到用遗传学方法研究细胞周期的可能性。他采用啤酒酵母细胞建立系统模型,经过一系列试验,分离出细胞周期基因发生突变的酵母细胞。Hartwell和其他科学家相继发现了100多种与细胞周期调控相关的CDC基因族。其中,Hartwell发现的CDC28调控细胞周期G1期进程的第一步,故又称为“start”基因。另外,Hartwell在研究酵母细胞对辐射的敏感性基础上,提出了著名的“checkpoint”概念,即当DNA受损时,细胞周期会停止。这一现象的生理意义在于,在细胞进入下一个细胞周期之前能有足够的时间进行DNA修复。后来,Hartwell将“checkpoint”的概念扩展到调控并保障细胞周期各期之间的正确顺序。 Sir Paul (1949年生)继Hartwell之后在70年代中期采用非渊粟酒裂殖酵母细胞为模型,发现了cdc2基因在细胞分裂(从G2期到有丝分裂期)调控方面起重要作用。后来,他发现cdc2与Hartwell在啤酒酵母中发现的“start”基因相同,还可调控从G1期到S期的转变。因此,cdc2基因可调控细胞周期的不同阶段。 1987年,Nurse分离出人类的相应基因——CDK1。Nurse发现CDK的活性依赖可逆性的磷酸化反应。基于这些理论,又有一些人类的CDK分子相继被发现。R. Timothy Hunt(1943年生)在80年代早期发现了第一个周期蛋白分子。周期蛋白是一种在细胞周期中周期性产生和降解的蛋白质。周期蛋白与CDK分子结合,调节CDK的活性。Hunt首先发现,在海胆细胞中周期蛋白在细胞周期中会发生周期性的降解,这是调控细胞周期的重要机制。Hunt在其他物种中也发现了周期蛋白,这些周期蛋白在进化过程中高度保守。 3位诺贝尔奖获得者创建了细胞周期调控的分子机制。CDK分子的含量在细胞周期中是恒定的,但是它的活性却因周期蛋白的调控作用而不同。周期蛋白和CDK分子共同驱动细
Section A 细胞与大分子 简述复杂大分子的生物学功能及与人类健康的关系。 Section C 核酸的性质 1.DNA的超螺旋结构的特点有哪些? A 发生在闭环双链DNA分子上 B DNA双链轴线高卷曲,与简单的环状相比,连接数发生变化 C 当DNA扭曲方向与双螺旋方向相同时,DNA变得紧绷,为正超螺旋,反之变得松弛为负超螺旋。自然界几乎所有DNA分子超螺旋都为负的,因为能量最低。 2.简述核酸的性质。 A 核酸的稳定性:由于核酸中碱基对的疏水效应以及电荷偶极作用而趋于稳定 B 酸效应:在强酸和高温条件下,核酸完全水解,而在稀酸条件下,DNA的核苷键被选择性地断裂生成脱嘌呤核酸 C 碱效应:当PH超出生理范围时(7-8),碱基的互变异构态发生变化 D 化学变性:一些化学物质如尿素,甲酰胺能破坏DNA和RNA二级结构中的 而使核酸变性。 E 粘性:DNA的粘性是由其形态决定的,DNA分子细长,称为高轴比,可被机械力和超声波剪切而粘性下降。 F 浮力密度:1.7g/cm^3,因此可利用高浓度分子质量的盐溶液进行纯化和分析 G 紫外线吸收:核酸中的芳香族碱基在269nm 处有最大光吸收 H 减色性,热变性,复性。 思考题:提取细菌的质粒依据是核酸的哪些性质? 质粒是抗性基因,,在基因组或者质粒DNA中用碱提取法。 Sectio C 课前提问 1.在1.5mL的离心管中有500μL,取出10 μL稀释至1000 μL后进行检测,测得A260=0.15。 问(1):试管中的DNA浓度是多少? 问(2):如果测得A280=0.078, .A260/A280=?说明什么问题? (1)稀释前的浓度:0.15/20=0.0075 稀释后的浓度:0.0075/100=0.75ug/ml (2)0.15/0.078=1.92〉1.8,说明DNA中混有RNA样品。 2.解释以下两幅图
分子生物学 第一章绪论 分子生物学研究内容有哪些方面? 1、结构分子生物学; 2、基因表达的调节与控制; 3、DNA重组技术及其应用; 4、结构基因组学、功能基因组学、生物信息学、系统生物学 第二章DNA and Chromosome 1、DNA的变性:在某些理化因素作用下,DNA双链解开成两条单链的过程。 2、DNA复性:变性DNA在适当条件下,分开的两条单链分子按照碱基互补原则重新恢复天然的双螺旋构象的现象。 3、Tm(熔链温度):DNA加热变性时,紫外吸收达到最大值的一半时的温度,即DNA分子内50%的双链结构被解开成单链分子时的温度) 4、退火:热变性的DNA经缓慢冷却后即可复性,称为退火 5、假基因:基因组中存在的一段与正常基因非常相似但不能表达的DNA序列。以Ψ来表示。 6、C值矛盾或C值悖论:C值的大小与生物的复杂度和进化的地位并不一致,称为C值矛盾或C值悖论(C-Value Paradox)。 7、转座:可移动因子介导的遗传物质的重排现象。 8、转座子:染色体、质粒或噬菌体上可以转移位置的遗传成分 9、DNA二级结构的特点:1)DNA分子是由两条相互平行的脱氧核苷酸长链盘绕而成;2)DNA分子中的脱氧核苷酸和磷酸交替连接,排在外侧,构成基本骨架,碱基排列在外侧;3)DNA分子表面有大沟和小沟;4)两条链间存在碱基互补,通过氢键连系,且A=T、G ≡ C(碱基互补原则);5)螺旋的螺距为3.4nm,直径为2nm,相邻两个碱基对之间的垂直距离为0.34nm,每圈螺旋包含10个碱基对;6)碱基平面与螺旋纵轴接近垂直,糖环平面接近平行 10、真核生物基因组结构:编码蛋白质或RNA的编码序列和非编码序列,包括编码区两侧的调控序列和编码序列间的间隔序列。 特点:1)真核基因组结构庞大哺乳类生物大于2X109bp;2)单顺反子(单顺反子:一个基因单独转录,一个基因一条mRNA,翻译成一条多肽链;)3)基因不连续性断裂基因(interrupted gene)、内含子(intron)、外显子(exon);4)非编码区较多,多于编码序列(9:1) 5)含有大量重复序列 11、Histon(组蛋白)特点:极端保守性、无组织特异性、氨基酸分布的不对称性、可修饰作用、富含Lys的H5 12、核小体组成:由组蛋白和200bp DNA组成 13、转座的机制:转座时发生的插入作用有一个普遍的特征,那就是受体分子中有一段很短的被称为靶序列的DNA会被复制,使插入的转座子位于两个重复的靶序列之间。 复制型转座:整个转座子被复制,所移动和转位的仅为原转座子的拷贝。 非复制型转座:原始转座子作为一个可移动的实体直接被移位。 第三章DNA Replication and repair 1、半保留复制:DNA生物合成时,母链DNA解开为两股单链,各自作为模板(template)按碱
细鳞斜颌鲴种群的遗传分化及系统发生生物地理学研究 武震M100102115水生生物学 摘要:细鳞斜颌鲴(Xenocypris microlepis)属鲤形目,鲤科,鲴亚科,鲴属。俗称:沙姑子、黄片。我们将以中国各水系细鳞斜颌鲴种群为研究对象,以基因组微卫星标记和线粒体D-loop标记为线索,研究细鳞斜颌鲴种群的遗传分化及系统发生生物地理学特征,探讨相互间的遗传结构、亲缘关系和系统进化关系,为进一步开发和利用细鳞斜颌鲴资源奠定基础。 关键字:细鳞斜颌鲴,线粒体D-loop标记,微卫星标记,遗传分化, 亲缘关系, 系统进化 1.研究背景 细鳞斜颌鲴属中下层鱼类,平时喜生活于江河干支流水域,到了产卵季节,有一定的短距离洄游现象,上溯至适合条件的产卵场进行集群产卵。产后,亲鱼分散游动,离开产卵场,至秋季有一部分群体进入干流附属的湖泊或支流中进行索饵、育肥,冬季则又返回干流水深的潭穴中越冬。细鳞斜颌鲴的食性很杂,自全长2厘米以上的夏花鱼种开始,除摄食少量浮游生物外,主要是腐屑、底泥以及底生硅藻和摇蚊幼虫等底生生物。它在不同类型的水体中,均以腐殖质有机碎屑、腐泥及着生藻类为主要食物。其生长在头两年速度较快,2龄鱼的平均体重可达479克。细鳞斜颌鲴通常2冬龄性成熟,生殖季节在华中和华南地区为4―6月。成熟雌鱼的体重变化在415―1100克以上。平均每千克体重的鱼怀卵量为20万粒左右。产粘性卵,呈浅黄色。产出时卵径为0.8―1.2毫米。雄鱼在生殖季节,有珠星出现。广泛存在于东部各水系之中。故各水系之间的种群长期存在地理隔离,基因交流困难,是一个良好的进化生态学研究材料。国内对此鱼的研究也不多,且多为形态学方面的资料,研究其分子进化和群体遗传,有助于了解该种的资源状况,同时能够为生态学相关理论提供依据。 2.方法 2.1采样 分别采钱塘江,长江,珠江水系细鳞斜颌鲴,每条水系定5—7个点,如钱
分子生物学总结完整版 1、结构分子生物学; 2、基因表达的调节与控制; 3、DNA重组技术及其应用; 4、结构基因组学、功能基因组学、生物信息学、系统生物学 第二章DNA and Chromosome 1、DNA的变性:在某些理化因素作用下,DNA双链解开成两条单链的过程。 2、 DNA复性:变性DNA在适当条件下,分开的两条单链分子按照碱基互补原则重新恢复天然的双螺旋构象的现象。 3、 Tm(熔链温度): DNA加热变性时,紫外吸收达到最大值的一半时的温度,即DNA分子内50%的双链结构被解开成单链分子时的温度) 4、退火:热变性的DNA经缓慢冷却后即可复性,称为退火 5、假基因:基因组中存在的一段与正常基因非常相似但不能表达的DNA序列。以Ψ来表示。 6、 C值矛盾或C值悖论:C值的大小与生物的复杂度和进化的地位并不一致,称为C值矛盾或C值悖论(C-Value Paradox)。 7、转座:可移动因子介导的遗传物质的重排现象。 8、转座子:染色体、质粒或噬菌体上可以转移位置的遗传成分
9、 DNA二级结构的特点:1)DNA分子是由两条相互平行的脱氧核苷酸长链盘绕而成;2)DNA分子中的脱氧核苷酸和磷酸交替连接,排在外侧,构成基本骨架,碱基排列在外侧;3)DNA分子表面有大沟和小沟;4)两条链间存在碱基互补,通过氢键连系,且A=T、G ≡ C(碱基互补原则);5)螺旋的螺距为 3、4nm,直径为2nm,相邻两个碱基对之间的垂直距离为0、34nm,每圈螺旋包含10个碱基对;6)碱基平面与螺旋纵轴接近垂直,糖环平面接近平行 10、真核生物基因组结构:编码蛋白质或RNA的编码序列和非编码序列,包括编码区两侧的调控序列和编码序列间的间隔序列。特点:1)真核基因组结构庞大哺乳类生物大于2X109bp;2)单顺反子(单顺反子:一个基因单独转录,一个基因一条mRNA,翻译成一条多肽链;)3)基因不连续性断裂基因(interrupted gene)、内含子(intron)、外显子(exon);4)非编码区较多,多于编码序列(9:1) 5)含有大量重复序列1 1、Histon(组蛋白)特点:极端保守性、无组织特异性、氨基酸分布的不对称性、可修饰作用、富含Lys的H5 12、核小体组成: 由组蛋白和200bp DNA组成 13、转座的机制:转座时发生的插入作用有一个普遍的特征,那就是受体分子中有一段很短的被称为靶序列的DNA会被复
第六章细胞周期及其调节 细胞增殖(cell proliferation)与细胞生长分裂周期. 第一节细胞周期 一、细胞周期(cell cycle):指亲代细胞分裂结束到子代细胞分裂结束所经历的过程,这个过程所需的时间称为细胞周期时间。 细胞周期由G1、S、G2和M期组成(G1、S和G2期又合称为分裂间期)。 G1(Gap1)期:DNA合成前期(复制前期),从上次有丝分裂完成到DNA复制之前的阶段; S期:DNA复制期; G2期:合成后期,从DNA复制完成至有丝分裂开始; M期:有丝分裂(Mitosis)期,包括核分裂和胞质分裂. M期结束后形成两个新的子细胞。 注:①不同细胞的细胞周期时间不同,一般S+G2+M期较恒定,而G1期变化较大,因而它决定了细胞周期时间的长短; ②G1期细胞有三种可能的趋向:1)进入S期(即进入细胞周期).2)处于静止期即Co期(在一定条件下可重新进入增殖周期),3)分化、衰老、凋亡。 二、细胞周期中各时相的主要生化事件 细胞周期中每期都有其特殊功能,其中S期的DNA复制和M期细胞核的有丝分裂是细胞周期中2个最关键的过程: 1、G1期:为DNA复制作准备,G1早期合成各种RNA、结构蛋白和酶等,细胞通过一 1
个限制点(restriction point,R点)后在G1后期合成DNA复制有关的蛋白和酶。 在开始合成DNA之前有一个关卡(checkpoint),检查染色体DNA是否有损伤,如有则先要进行修复。 2、S期:DNA(包栝端粒)的复制及组蛋白合成、核小体装配.S期后每一染色体复制成2个染色单体· S→G2期关卡:检查DNA复制是否完成 3、G2期:为有丝分裂作准备.有RNA和非组蛋白合成。 4、M期:染色体浓缩一仿锤体形成→染色体分离并移向细胞两端→染色体解聚,形成两个新核→胞质分裂。 第二节周期素依赖性蛋白激晦与细胞周期调节 周期素依赖性蛋白激酶(cyclin-dependent kinases,CDKs) 通过使特异底物磷酸化调节细胞周期进行,其活性依赖与周期素(cyclin)结合形成复合物。 一、周期素-周期素依赖性蛋白激酶 周期素家族和周期素依赖蛋白激酶(CDK)家族. 细胞周期的不同时相表达不同cyc-CDK,这些cyc-CDK复合物在各不同的细胞周期过渡点起作用. 1、G1期cyc-CDK G1期表达的周期素为周期素C、D(D1、D2、D3)和E。 D族周期素主要与CDK4(以及CDK2、CDK5、CDK6)结合成活性的蛋白激酶复合物,对细胞通过R点(G0→G1过渡有重要作用。 E族周期素与CDK2形成复合物。 cycE-CDK2复合物调控G1→S过渡。 2
细胞和分子生物学实验重点知识点汇总 Experiment1细胞有丝分裂 间期:有明显的细胞核,染色质分布较均均,由于染色质易与碱性染料结合,故细胞核的染色比细胞质深。核中可见1~3个染色较浅的呈球状的核仁 前期:细胞核膨大,染色质逐渐螺旋化为丝状的染色丝,其后染色丝进一步缩短变粗,形成一定形态和书目的染色体(这时候的每条染色体由两条染色单体组成,但在光镜下一般不易看清),核膜、核仁逐渐消失 中期:每条染色体中的成对染色单体逐渐分开(但着丝粒仍未分离)全部染色体(2n=16)移向细胞中央的赤道面上,形成赤道板。在赤道板到两面有许多纺锤丝连接细胞两极和染色体的着丝点,成为纺锤体,但不易观察到,此时染色体形态最典型 后期:着丝粒纵裂为二。这是,每条染色体的两条染色单体已完全分开,由于纺锤丝的牵引,分别向细胞的两极移动,形成了数目相等的两组染色体(这是所观察到的染色体数目比原来增加1倍,是由于S期内DNA含量倍增的结果) 末期:染色体移到两极并解旋为染色质,细胞中部出现细胞板,并逐渐向边缘发展。当染色质构成核网时,核膜、核仁重新出现。细胞板达到两边,分裂结束,形成两个子细胞,细胞又进入间期状态。 Experiment2动物染色体的制备 原理:染色体只有在分裂期的细胞,特别是中期细胞中表现出典型形态便于观察和计数,所以必须采取特殊的技术方法,从发生有丝分裂的组织和细胞悬液中得到。最常用的途径是从骨髓细胞、血淋巴细胞和组织培养的细胞中制备。骨髓细胞数量多、分裂旺盛,不需体外培养和无菌操作,便于取材。 秋水仙素的作用:抑制纺锤体的形成,使细胞停留在分裂中期 KCl低渗溶液:使细胞膨胀,促使中期染色体散开 固定液:有固定作用,对染色体还有一定的分散作用 Giemsa染色液:染色 结果:低倍镜下,可见到许多大笑不等被染成紫红色呈圆形的间期细胞核以及分散在它们之间的中期分裂象。小鼠染色体一般呈“U”形,染色体2n=40
分子生物学常见名词解释完全版(中英文对照) A Abundance (mRNA 丰度):指每个细胞中mRNA 分子的数目。 Abundant mRNA(高丰度mRNA):由少量不同种类mRNA组成,每一种在细胞中出现大量 拷贝。 Acceptor splicing site (受体剪切位点):内含子右末端和相邻外显子左末端的边界。Acentric fragment(无着丝粒片段):(由打断产生的)染色体无着丝粒片段缺少中心粒,从而 在细胞分化中被丢失。 Active site(活性位点):蛋白质上一个底物结合的有限区域。 Allele(等位基因):在染色体上占据给定位点基因的不同形式。 Allelic exclusion(等位基因排斥):形容在特殊淋巴细胞中只有一个等位基因来表达编码的 免疫球蛋白质。 Allosteric control(别构调控):指蛋白质一个位点上的反应能够影响另一个位点活性的能力。Alu-equivalent family(Alu 相当序列基因):哺乳动物基因组上一组序列,它们与人类Alu 家族相关。 Alu family (Alu家族):人类基因组中一系列分散的相关序列,每个约300bp长。每个成员 其两端有Alu 切割位点(名字的由来)。 α-Amanitin(鹅膏覃碱):是来自毒蘑菇Amanita phalloides 二环八肽,能抑制真核RNA聚 合酶,特别是聚合酶II 转录。 Amber codon (琥珀密码子):核苷酸三联体UAG,引起蛋白质合成终止的三个密码子之一。Amber mutation (琥珀突变):指代表蛋白质中氨基酸密码子占据的位点上突变成琥珀密码 子的任何DNA 改变。 Amber suppressors (琥珀抑制子):编码tRNA的基因突变使其反密码子被改变,从而能识 别UAG 密码子和之前的密码子。 Aminoacyl-tRNA (氨酰-tRNA):是携带氨基酸的转运RNA,共价连接位在氨基酸的NH2 基团和tRNA 终止碱基的3¢或者2¢-OH 基团上。 Aminoacyl-tRNA synthetases (氨酰-tRNA 合成酶):催化氨基酸与tRNA 3¢或者2¢-OH基团共价连接的酶。 Amphipathic structure(两亲结构):具有两个表面,一个亲水,一个疏水。脂类是两亲结构,一个蛋白质结构域能够形成两亲螺旋,拥有一个带电的表面和中性表面。 Amplification (扩增):指产生一个染色体序列额外拷贝,以染色体内或者染色体外DNA形 式簇存在。 Anchorage dependence (贴壁依赖):指正常的真核细胞需要吸附表面才能在培养基上生长。Aneuploid (非整倍体):组成与通常的多倍体结构不同,染色体或者染色体片段或成倍丢失。Annealing (退火):两条互补单链配对形成双螺旋结构。 Anterograde (顺式转运):蛋白质质从内质网沿着高尔基体向质膜转运。 Antibody (抗体):由B 淋巴细胞产生的蛋白质(免疫球蛋白质),它能识别特殊的外源“抗 2 原”,从而引起免疫应答。 Anticoding strand (反编码链):DNA 双链中作为膜板指导与之互补的RNA 合成的链。Antigen (抗原):进入基体后能引起抗体(免疫球蛋白质)合成的分子。 Antiparallel (反式平行):DNA双螺旋以相反的方向组织,因此一条链的5¢端与另一条链的3¢端相连。
浅谈细胞周期调控 朱春森 摘要:近年来有关细胞周期调控机制研究进展较快,细胞周期调控可分为G1期调控和非G1期调控。在G1期调控中,细胞周期蛋白依赖性激酶复合体CDK激活后,通过Rb蛋白和转录因子启动基因转录。P16、p21、p15等蛋白通过抑制CDK的活性而发挥作用。P53蛋白和mdm2蛋白协同调节细胞周期活动。细胞周期的停滞或细胞凋亡对维护基因组稳定有重要意义。 关键词:细胞周期调控 Cyclin CDK CDI 调控机制 细胞周期调控是指各种调控因子通过自身的激活和灭活,使细胞启动和完成细胞周期重要事件,并保障这些事件按次序正常进行。细胞周期调控对维护基因组的稳定有着重要的意义。 1. 细胞周期调控的分子基础 细胞周期调控的分子基础包括细胞周期蛋白(Cyclin)、细胞周期蛋白依赖蛋白激酶(CDK)和细胞周期蛋白依赖蛋白激酶抑制物(CDI)。它们分别包括CyclinA、CDK17和p21、p27、p18等,p53和视网膜母细胞瘤蛋白(pRb)也参与细胞周期调控。 Cyclin 周期蛋白不仅仅起激活CDK的作用,还决定了CDK何时、何处、将何种底物磷酸化,从而推动细胞周期的前进。目前从芽殖酵母、裂殖酵母和各类动物中分离出的周期蛋白有30余种,在 脊椎动物中为A 1-2、B 1-3 、C、 D 1-3 、E 1-2 、F、G、H等。分为G 1 型、G 1 /S型S型和M型4类(见表 1)。各类周期蛋白均含有一段约100个氨基酸的保守序列,称为周期蛋白框,介导周期蛋白与CDK结合。 表1不同类型的周期蛋白 *包括D1-3,各亚型cyclin D,在不同细胞中的表达量不同,但具有相同的功效 CDK
, 内膜系统的膜结构破裂后自己重新封闭起来的小囊泡(主要 是内质网和高尔基体), 是异质性的集合体, 形态、大小及功能常因生物种类和细胞类型不同而异。据微体内含有的酶的不同可分为过氧化物酶体、糖酵解酶体和乙醛酸循环体。在蛋白质合成过程中,同一条mRNA分子能够同多个核糖体结合,同时合成若干条蛋白质多肽链,结合在同一条mRNA上的核糖 叠的多肽链相互作用的蛋白质,能够加速正确折叠的进行或提供折叠发生所需要的微环境。动物体细胞在体外可传代的次数,与物种的寿命有关,它们的增殖能力不是无限的, DNA在核小体连接处断裂成核小体片 色体末端的特殊结构,即染色体末端DNA 序列的多个重复,其作用是保护和稳定染色 RNA 依赖性DNA 聚合酶,为一种核糖核蛋白酶,是合成端粒必需的酶。在双线期中,交叉数目逐渐减少,在着丝粒两侧的交叉向两端移动.这个现象称为 成染色体联会的两条同源染色体互相紧靠,进而缠绕在一起,基质开始附着到染色丝上,成为一条短而粗的染色体。据染色体被拉向两极所受到的力的不同,后期可分为后期A 和后期B,此时的染色体 启动DNA复制的关键因子,是真核细胞DNA M期促进因子。
能够促使染色体凝集,使细胞由G2期进入M 物质多肽的形式合成,其N末端含有作为通过膜时之信号的氨基酸序列。引导前体多肽 是指具有摄取、处理及提呈抗原能力的细胞,能摄取病原体蛋白并将其加工将成短肽段,呈递给T细胞。 ,从中 于高等真核细胞中,是内层核被膜下纤维蛋白片层,纤维纵横排列整齐呈纤维网络状。 成串排列在一起,主要集中在染色体的着丝 DNA和组蛋白构成,是染色质的基本结构 在一定时期的特种细胞的细胞核内, 它由不表达的DNA序列组成, 分裂过程中,核仁出现周期性变化。一般在分裂前期逐渐消失,其纤丝和颗粒成分散失于核质之中;在分裂末期又重新出现。核仁的形成常与特定染色体的一定区域密切相关。 色体片段, 通过次缢痕与染色体主要部分相连。 指染色体组在有丝分裂中期的表型, 是染色体数目、大小、 是卵母细胞进行第一次减数分裂时, 停留在双线期的染色体。含4条染色单体,形似灯刷。 由核内有丝分裂产生的多股染色单体平行排列而成。
名词解释 1. 基因(gene): 2. 结构基因(structural gene): 3. 断裂基因(split gene): 4. 外显子(exon): 5. 内含子(intron): 6. 多顺反子RNA(polycistronic/multicistronic RNA): 7. 单顺反子RNA(monocistronic RNA): 8. 核不均一RNA(heterogeneous nuclear RNA, hnRNA): 9. 开放阅读框(open reading frame, ORF): 10. 密码子(codon): 11. 反密码子(anticodon): 12. 顺式作用元件(cis-acting element): 13. 启动子(promoter): 14. 增强子(enhancer): 15. 核酶(ribozyme) 16. 核内小分子RNA(small nuclear RNA, snRNA) 17. 信号识别颗粒(signal recognition particle, SRP) 18. 上游启动子元件(upstream promoter element) 19. 同义突变(same sense mutation) 20. 错义突变(missense mutation) 21. 无义突变(nonsense mutation) 22. 移码突变(frame-shifting mutation) 23. 转换(transition) 24. 颠换(transversion) (三)简答题 1. 顺式作用元件如何发挥转录调控作用? 2. 比较原核细胞和真核细胞mRNA的异同。 3. 说明tRNA分子的结构特点及其与功能的关系。 4. 如何认识和利用核酶? 5. 若某一基因的外显子发生一处颠换,对该基因表达产物的结构和功能有什么影响? 6. 举例说明基因突变如何导致疾病。 (四)论述题 1. 真核生物基因中的非编码序列有何意义? 2. 比较一般的真核生物基因与其转录初级产物、转录成熟产物的异同之处。 3. 真核生物的基因发生突变可能产生哪些效应? (二)名词解释 1.基因组(genome) 2. 质粒(plasmid) 3.内含子(intron) 4.外显子(exon) 5.断裂基因(split gene) 6.假基因(pseudogene) 7.单顺反子RNA(monocistronic RNA)
细胞周期的关键分子调节机制 摘要:细胞周期是指连续分裂的细胞从一次有丝分裂结束到下一次有丝分裂完成所经历的整个序贯过程. 细胞周期中每一事件都是有规律、精确地发生, 并且在时间与空间上受到严格调控. 细胞周期中最关键的三类调控因子是: cdc 基因、周期蛋白依赖性激酶( CDKs) 及细胞周期蛋白( cyclin) . 这些调控因子的发现对肿瘤学及发育生物学的发展都有重要的理论和实践意义. 关键词:细胞周期, cdc 基因, 周期蛋白依赖性激酶, 细胞周期蛋白 细胞周期( cell cycle) 是保证细胞正确增殖的过程, 对一个细胞而言, 在分裂过程中获得生存所必需的物质是最关键的环节, 尤其是合成遗传所需物质 . 细胞周期可划分为4 个时相, 即G1、S、G2 和M 期. 在G1 期中, 细胞不断生长发育. 当达到一定体积时, 细胞就会进入DNA 合成( S)期, 细胞内遗传物质开 始复制, 最终形成两套完整的染色体组( chromosome set) , 细胞便进入有丝分裂前的准备( G2) 期. 在有丝分裂(M) 期, 染色体组分离、细胞质分裂, 两套染色体平均分配给两个子细胞, 从而完成一个细胞周期( 图1) . 图1 细胞周期4 个时相 细胞周期中最关键的三类调控因子是: cdc 基因、周期蛋白依赖性激酶( CDKs) 及细胞周期蛋白( cyclin) . 1.cdc基因
1.1 cdc基因的发现 哈特韦尔采用遗传学方法, 用芽殖酵母( Saccha rymyces cerevisiae)作为实验对象研究细胞周期.20世纪70年代初,他通过温度敏感突变技术筛选出突变酵母细胞, 这些细胞的生长停滞在特定的细胞周期时相 , 从而确定缺陷基因所编码的蛋白质在细胞周期调控中的作用,利用种方法,他成功地分离出上百个涉及细胞周期调控的基因(图2),并命名为cdc 基因. 图2 用荧光钙( calcofluor) 示裂殖酵母(Schizosacchharomyces pombe) 细胞壁和中隔( septum) 野生型细胞的长度加倍并一分为二, 而cdc25 缺陷的细胞已长的很长却不分裂. cdc25 是细胞从G2 期进入M 期必需的基因, 它负责CDK2 的去磷酸化( 引自Nurse P) . 1.2 cdc基因的功能 在哈特韦尔发现的这类基因中, cdc4、6、7、8 等控制DNA 复制, 如cdc8 具有起始DNA 合成的功能 ; cdc5、14、15 等参与染色体分离的调控; cdc3、10、11、13 等调控细胞质的分裂,名为cdc28 的基因, 启动细胞从G1 期进入S期. 该基因编码的蛋白质是其他cdc 基因产物执行功能的前提, 所以又被称为star t基因。 2.周期蛋白依赖性激酶(CDK)
细胞与分子生物学考题 Chapter 3 Protein Structure & Function 1. The primary, secondary, tertiary and quaternary structures of proteins. N972010028 黄琴淑 (1) 一级结构 (primary structure) :蛋白质的序列称之为蛋白质的「一级结构」。 (2) 二级结构 (secondary structure) : 一级结构上的胺基酸间可交互作用,利用醯胺键上的C=O键与胺基形成氢键。这样形成的简单又有规则的结构,称之为二级结构 (secondary structure)。蛋白质有α螺旋 (helix)与 beta 折曲平面 (pleated sheet); 两种主要 而且规则的二级结构,由这些简单的结构又可组合成一些独立折叠的单元,称之为模组(motif)。 (3) 三级结构 (tertiary structure) :蛋白质的三级结构是由一条多月生(polypeptide)链组成,可包含一个或多个模组。 (4) 四级结构 (quaternary structure):蛋白质的三级结构是由一条多月生(polypeptide)链组成,可包含一个或多个模组。一个含有多个次单元蛋白质中,每个次单元都是一个三级结构,次单元间可能有疏水性作用,盐桥等交互作用而形成四级结构,所以含有多个次单元的蛋白质才有四级结构 (quaternary structure)。 第壹题参考资料 蛋白质的一级结构 将蛋白质中胺基酸顺序视为整体构造,是一种用有机化学词语来描述分子的完全方法。自很多不同蛋白质的顺序分析中可以看出,每种蛋白质都有其独特的结构,而顺序排列即是该种系的特性。在少数的情形中,特殊的器官或组织也具有特定结构的蛋白质,更进一步的,蛋白质可以如细胞分裂一般很正确地被复制出相同顺序的蛋白质。我们可以参考牛的胰岛素(Bovine insulin);更正确地说,是参考proinsulin。Proinsulin为生物活性贺尔蒙的先质(precursor),藉着正常牛的胰脏岛状细胞仔,细地做成的一种特定构造。牛的胰岛素和其他哺乳类的胰岛素几乎是相同的,通常只有一个胺基酸不同,即A链中第8,9或10位置胺基酸的改变。这些相似性使得这方面的研究迅速扩展,虽然在不同种类中,胺基
《生物工程进展》1999,V ol.19,No.4 细胞周期检查点的分子机制 吴家睿 (中国科学院上海生物化学研究所 上海 200031) 摘要 细胞在长期的进化过程中发展出了一套保证细胞周期中DNA复制和染色体分配质量的检查机制,通常被称为细胞周期检查点(checkpoint)。当细胞周期进程中出现异常事件,这类调节机制就被激活,及时地中断细胞周期的运行,待细胞修复或排除了故障后,细胞周期才能恢复运转。根据“质量控制”的内容,可将细胞周期检查点分为三种。第一种负责查看DNA有无损伤,称为DNA损伤检查点;第二种负责DNA复制的进度,称为DNA复制检查点;第三类是管理染色体的正确分配与否,称为纺锤体组装检查点。本文围绕检查点工作的这三个方面对其分子机制作一个简单的论述。 关键词 细胞周期 检查点 DN A复制 DNA损伤 染色体分配 细胞周期最主要的任务是将其基因组DNA在DNA合成期(S期)完整地复制成两份拷贝,然后在分裂期(M期)将这两份拷贝正确无误地分配给两个子代细胞。如果在这一任务的完成过程中产生的错误没有得到及时纠正,那么将导致基因组的不稳定和变异。对单细胞生物而言,其后果是导致细胞增殖能力下降或细胞死亡。对多细胞生物而言则通常是引起肿瘤。细胞在长期的进化过程中发展出了一套保证细胞周期中DNA复制和染色体分配质量的检查机制,通常被称为细胞周期检查点(check-point)。这是一类负反馈调节机制。当细胞周期进程中出现异常事件,如DNA损伤或DNA复制受阻,这类调节机制就被激活,及时地中断细胞周期的运行,待细胞修复或排除了故障后,细胞周期才能恢复运转。 根据“质量控制”的内容,可将细胞周期检查点分为三种。第一种负责查看DNA有无损伤,称为DNA损伤检查点(DNA dam ag e checkpo int);第二种负责DN A复制的进度,称为DNA复制检查点(DNA r eplication check-point);第三类是管理染色体的正确分配与否,称为纺锤体组装检查点(Spindle assemmbly checkpo int),因为染色体的分配主要依赖于纺锤体的作用。近几年来,在检查点如何工作的分子机制的研究方面已有了许多进展。本文试图围绕检查点工作的这三个方面作一个简单的论述。 1 DNA损伤检查点 检查点工作的第一步是发现问题。一般认为,DNA损伤的出现可以迅速地激活DNA损伤检查点。由于DNA损伤可以发生在细胞周期的任一个时期,包括G1期、S期、G2/M期中,且DNA损伤有许多种类,所以存在多种探测DNA损伤的手段。它们可以在不同的时期对特定的DNA损伤进行检测。在酵母细胞中, DNA链的断裂和单链DNA的形成可以产生损伤起始信号,由至少两类检查点蛋白进行感知。一类是Rad9;另一类是Rad24,Rad17, M ec3和Ddc1。最新的研究发现,Rad9蛋白在DNA损伤的情况下,被M ec1和T el1激酶磷酸化,然后与Rad53蛋白结合[1]。研究者通过双杂交和免疫共沉淀等方法揭示,Rad17,M ec3和Ddc1形成蛋白复合体,其功能受Rad24的调节,即位于Rad24的下游[2]。Rad24能与DNA复制的辅助蛋白RFC上的亚基Rfc2结合[3]。因此,Rad24可能与RFC形成探测器,一 7
802细胞与分子生物学考试大纲(2015版) 细胞生物学部分 1 绪论 细胞生物学的主要研究内容与当前细胞生物学研究的根本问题,细胞学说的创立及其内容要点与意义。 2 细胞的统一性与多样性 细胞的基本特征,原核细胞与古核细胞、真核细胞以及非细胞生命体的基本知识。 3、细胞生物学研究方法 细胞形态结构的观察方法与相关仪器的原理与应用范围,细胞化学组成及其定位与动态分析技术的原理与应用范围,细胞培养及细胞工程的相关概念与方法原理,细胞及生物大分子动态变化研究方法的概念及原理,细胞生物学研究中常用的模式生物,功能基因组学的基本研究思路与方法。 4、细胞质膜 细胞质膜结构模型的基本要点,细胞质膜的基本组成成分及其特点与意义,细胞质膜的基本特征、功能与研究方法。 5、物质的跨膜运输 物质的跨膜运输的基本概念,跨膜运输的主要途径、转运装置、运输的基本过程。 6、线粒体与叶绿体。 线粒体的基本形态,动态特征及其分子细胞生物学基础,线粒体超微结构组成及其功能特点,氧化磷酸化的分子结构基础与转化机制,线粒体的半自主性与起源。 7、细胞质基质与细胞内膜系统 细胞质基质的含义与功能。 内膜系统的概念及其组成成员;内质网的基本类型及其功能,内质网应激及其信号调控;高尔基复合体的形态结构、标志性酶以及功能;溶酶体与过氧化物酶体的结构特点,发生与功能。
8、蛋白质分选与膜泡运输 信号假说与蛋白质分选信号。蛋白质分选的基本途径与类型。蛋白质向线粒体与过氧化物酶体的分选途径与机制。膜泡运输的途径与机制,细胞结构体系的组装方式及意义。 9、细胞信号转导 细胞信号转导的基本知识与基本概念,各种类型信号传递的通路,细胞信号转导的整合与控制。 10、细胞骨架 细胞骨架的基本概念。 微丝的组成及其组装,网格结构的调节与细胞运动,依赖于微丝的分子马达,以及肌细胞收缩运动结构基础与机制模型;微管的结构组成及其极性,组装与去组装,微管组织中心,微管的动力学性质,微管网格结构的调节,微管的功能(包括对细胞结构的组织作用,物质运输,纤毛与鞭毛的结构与功能,纺锤体);中间丝的一般形态与类型及其细胞特异性,中间丝的组装与表达,中间丝与其她细胞结构的联系。 11、细胞核与染色体 核被膜的结构特点、崩解与组装、生物学意义;核孔复合体的结构模型及功能;核纤层的蛋白组成与功能。染色质的概念及其化学组成,基因组DNA的类型,染色质蛋白的的类型与特性;核小体的发现与结构;染色质的组装;染色质的类型及其特性。染色质复制与修复、表达的基本概念与调控机制。染色体的形态结构及其相关概念,染色体DNA的功能元件。核仁的超微结构分部与各部分的结构组成特点,核仁的功能,核仁周期性。 12、核糖体 核糖体的结构成分及其功能,核糖体的本质,RNA在生命起源中的作用。13、细胞周期与细胞分裂 细胞周期与分裂的相关的基本概念;细胞周期的时相划分及各时相的主要事件,以及研究细胞周期的最基本方法,早期胚胎与细菌细胞周期的特点。细胞有丝分裂的形态学过程,时相划分及各时相的变化标志,早中期染色体的移动与纺锤体的形成与结构,姐妹着丝粒的分离与后期染色体的移动,胞质分裂;减数分裂的形态学过程,时期划分与各期的主要变化特征,重要事件,特殊结构及其变化。
附录常见细胞分子生物学名词及其释义 α-actinin α-辅肌动蛋白一种使肌动蛋白成束的蛋白,有两个相距较远的肌动蛋白结合位点,故形成的肌动蛋白纤维束较为松散。 Akinase (PKA) A激酶因细胞内cAMP浓度升高而被激活催化靶蛋白磷酸化的酶。accessorycell 辅佐细胞在免疫应答过程中,能摄取、加工、处理并将抗原信息提呈给淋巴细胞的免疫细胞,又称抗原提呈细胞. actin 肌动蛋白真核细胞中含量丰富,是构成肌动蛋白丝的一种蛋白质。单体称球形肌动蛋白(G-actin),聚合物称丝状肌动蛋白(F-actin)。 actin-bindingprotein 肌动蛋白结合蛋白在细胞中与肌动蛋白单体或肌动蛋白纤维结合的、能改变其特性的蛋白质。 actinin 辅肌动蛋白一种肌动蛋白结合蛋白,集中分布在Z线和与质膜结合的应力纤维点状黏附端。 actin-relatedprotein(ARP) 肌动蛋白相关蛋白促进肌动蛋白丝集结的蛋白质复合物。activetransport 主动运输溶质通过细胞膜逆浓度梯度运输的现象,是一个耗能的生理过程。 actomere 肌动蛋白粒由未聚合的抑丝蛋白—肌动蛋白复合物和一小段肌动蛋白丝束组成的结构。一旦抑丝蛋白—肌动蛋白复合物发生解离,则引起肌动蛋白聚合成丝。actomyosin 肌动球蛋白肌肉收缩时肌动蛋白与肌球蛋白瞬时接触形成的复合物。adaptin 衔接蛋白参与成笼蛋白衣被形成的一类蛋白质,能同时与跨膜受体以及成笼蛋白结合,在两者间起衔接作用。 adaptorprotein 衔接器蛋白在细胞内信号传递途径中,凡是在不同蛋白质问起连接作用的蛋白质的通称。 adducin 聚拢蛋白质膜骨架蛋白,为异二聚体。在钙离子浓度为毫摩级时,加速血影蛋白到血影蛋白—肌动蛋白复合物的装配。 adherensjunction 黏台连接在质膜的胞质面附着有肌动蛋白纤维的细胞连接,包括连接相邻的上皮细胞的黏着带和体外培养的成纤维细胞底面的黏着斑(focalcontact)。 adhesion plaque(focal adhesion,focal contact) 鞘着斑(斑状黏附) 细胞与非细胞性基底物间形成的黏附结构。该处的质膜中含有整联蛋白分子群,分子的胞外结构域与细胞外基质组分相连,胞内结构域通过接合器蛋白与微丝相连。 adhesion protein 黏附蛋白质存在于细胞外基质中的与细胞黏附于基质有关的一类蛋白质,包括纤连蛋白、层连蛋白和血纤蛋白原等。在细胞的黏附、迁移、增殖、分化等活动中起作用。 adult stem cell 成体干细胞;组织细胞(tissue stemcell) 存在于一种组织或器官分化细胞中的未分化细胞,具有自我更新的能力,并能分化成来源组织的主要类型特化细胞。有的成体干细胞具有可塑性,在一定条件下,可分化成许多不同类型的细胞。 allosome,heterochromosome 异染色体主要和全部由异染色质组成的染色体,如人的Y 染色体和超数B染色体。 amitosis 无丝分裂又称直接分裂,不形成染色体和纺锤体,细胞核直接一分为二,随后细胞质分裂成两个子细胞。多见于某些原生生物中,如纤毛虫等。 mnmlytical cytology 分析细胞学对细胞成分进行定性、定量研究的一门科学。 mphase 后期有丝分裂(或减数分数)过程中的一个阶段.在此阶段中姊妹染色单体分离,并向细胞两极移动,纺锤体延伸和纺锤体两极间距离增加。 anchorage-dependentcell (依赖)贴壁细胞只有贴附于不起化学作用的物体表面时才能生