第八章 神经胚和三胚层分化
- 格式:doc
- 大小:23.00 KB
- 文档页数:4

概述三胚层分化引言三胚层分化是胚胎发育过程中的一个重要阶段,指的是受精卵经过一系列细胞分裂和特定细胞移动后,形成三个不同的胚层:外胚层、中胚层和内胚层。
这三个胚层将进一步发展成为不同组织和器官,构成成熟的多细胞生物体。
胚胎发育过程1.受精卵:受精卵是由卵子和精子结合而成的单细胞体。
在受精卵形成后,它会经历一系列细胞分裂。
2.2细胞期:在受精卵内部,原核质被分割成两个相等大小的细胞。
3.4细胞期:原核质再次分割,形成四个相等大小的细胞。
4.8细胞期:原核质进一步分裂,生成八个相等大小的细胞。
5.胚泡期:在这一阶段,八个细胞开始紧密地排列在一起,并形成一个囊状结构,称为囊泡。
6.悬浮期:囊泡内细胞进一步分裂,增加细胞数目,但细胞大小并未改变。
7.囊胚期:在这一阶段,囊泡内的细胞开始分化成不同的层次,形成外胚层、中胚层和内胚层。
三胚层的形成1.外胚层:也称为表皮原肠。
它是由囊胚期的表面细胞发育而来。
外胚层会形成皮肤、毛发、指甲等表皮组织。
2.中胚层:也称为中间原肠。
它是由囊胚期的中间细胞发育而来。
中胚层会形成骨骼、肌肉、心脏等内部组织。
3.内胚层:也称为内部原肠。
它是由囊胚期的内部细胞发育而来。
内胚层会形成消化系统、呼吸系统、神经系统等内脏组织。
三个胚层的命运1.外胚层:外胚层最早分化出来,它将发展成为表皮和相关附件,如毛发和指甲。
外胚层还会分化成神经外胚层,形成神经系统的一部分。
外胚层还可以分化成生殖细胞,最终形成生殖器官。
2.中胚层:中胚层将发展成为骨骼、肌肉、心脏等内脏组织。
它还会分化成血液和血管系统的一部分。
3.内胚层:内胚层将发展成为消化系统、呼吸系统、泌尿系统和生殖系统等内脏组织。
它还会分化成神经系统的一部分。
调控三胚层分化的因素1.信号分子:在三胚层分化过程中,许多信号分子起着关键作用。
Wnt信号通路参与调控中胚层和内胚层的形成,而Fgf信号通路参与调控外胚层的形成。
2.转录因子:转录因子是调控基因表达的关键蛋白质。


概述三胚层分化1. 介绍三胚层分化是胚胎发育的一个重要过程,指的是胚胎中的细胞按照不同功能和组织的需求逐渐分化成不同的胚层。
这一过程发生在受精卵着床后的第三周左右,是胚胎发育的重要里程碑之一。
三胚层分化的过程涉及多个信号通路和分子机制的调控,对于胚胎发育和器官形成具有至关重要的作用。
2. 第一层胚层:外胚层外胚层是三胚层分化的第一层胚层,也称为原基胚层。
外胚层位于胚胎内侧,靠近子宫壁。
它主要分化为胚胎的皮肤、毛发、指甲等表皮组织,以及部分中枢神经系统和周围神经系统。
外胚层的分化是由一系列细胞信号通路和转录因子的调控完成的。
外胚层分化的过程中,细胞逐渐形成上皮细胞,并分化为外层的表皮细胞和内层的神经上皮细胞。
外层的表皮细胞进一步分化为表皮源性细胞和神经源性细胞,分别形成胚胎的皮肤和中枢神经系统。
内层的神经上皮细胞则会分化为神经元、胶质细胞等组织和细胞类型。
3. 第二层胚层:中胚层中胚层是三胚层分化的第二层胚层,也称为间基胚层。
中胚层位于外胚层和内胚层之间,是胚胎的中间层。
中胚层主要分化为胚胎的骨骼、肌肉、血液和泌尿系统等组织。
中胚层的分化过程中,细胞经历了多次迁移和重排,形成了各种不同的细胞类型和组织。
这一过程主要受到信号通路的调控,如Wnt信号通路、Fgf信号通路等。
通过这些信号通路的调节,细胞在中胚层中相互作用,定位,并逐渐分化为不同的组织和器官。
4. 第三层胚层:内胚层内胚层是三胚层分化的第三层胚层,也称为内基胚层。
内胚层位于胚胎的内部,靠近羊膜腔。
内胚层主要分化为胚胎的消化系统、呼吸系统、循环系统和生殖系统等组织。
内胚层的分化过程中,细胞逐渐形成上皮细胞并分化为不同的细胞类型。
这个过程中涉及到多个转录因子和信号通路的调控,如TGF-beta信号通路、Nodal信号通路等。
通过这些调控机制,内胚层的细胞定位并分化为消化系统的细胞、呼吸系统的细胞、循环系统的细胞和生殖系统的细胞等。
5. 总结三胚层分化是胚胎发育的一个关键过程,其中的外胚层、中胚层和内胚层分别分化为不同的组织和器官。

发育生物学试题库(发育生物学教学组)目录:第一章章节知识点与重点 (1)第二章发育生物学试题总汇 (6)第三章试题参考答案 (18)第一章章节知识点与重点绪论1.发育和发育生物学2.发育的功能3.发育生物学的基础4.动物发育的主要特点5.胚胎发育的类型(嵌合型、调整型)6.研究发育生物学的主要方法第一章细胞命运的决定1.细胞分化2.细胞定型及其时相(特化、决定)3.细胞定型的两种方式与其特点(自主特化、有条件特化)4.胚胎发育的两种方式与其特点(镶嵌型发育依赖型发育)5.形态决定子6.胞质定域(海胆、软体动物、线虫)7.形态决定子的性质8.细胞命运渐进特化的系列实验9.双梯度模型10.诱导11.胚胎诱导第二章细胞分化的分子机制1.细胞表型分类2.差异基因表达的源由3.了解基因表达各水平的一般调控机制第三章转录后的调控1.RNA加工水平调控2.翻译和翻译后水平调控第四章发育中的信号传导1.信号传导2.了解参与早期胚胎发育的细胞外信号传导途径第五章受精的机制1.受精2.受精的主要过程及相关知识3.向化性4.顶体反应5.皮质反应第六章卵裂1.卵裂特点(课堂作业)2.卵裂方式3.两栖类、哺乳类、鱼类、昆虫的卵裂过程及特点4.(果蝇)卵裂的调控机制第七章原肠作用1.了解原肠作用的方式:2.海胆、文昌鱼、鱼类、两栖类、鸟类、哺乳类的原肠作用基本过程与特点第八章神经胚和三胚层分化1.三个胚层的发育命运第九章胚胎细胞相互作用-诱导1.胚胎诱导和自动神经化、自动中胚层化2.胚胎诱导、异源诱导者3.初级诱导和次级诱导、三(多)级诱导4.邻近组织相互作用的两种类型5.间质与上皮(腺上皮)的相互作用及机制第十章胚轴形成1.体形模式2.图式形成3.果蝇形体模式建立过程中沿前后轴不同层次基因的表达4.果蝇前后轴建立的分子机制5.果蝇背腹轴形成的分子机制第十一章脊椎动物胚轴的形成1.什么是胚轴2.两栖类胚轴形成过程及分子机制。

![[知识]三个胚层的分化过程](https://uimg.taocdn.com/3fc4bc0e2379168884868762caaedd3383c4b564.webp)
三个胚层的分化过程三个胚层的分化过程:1.外胚层的分化脊索形成后,诱导其背侧中线的外胚层增厚呈板状,称神经板(neural plate)。
神经板随脊索的生长而增长,且头侧宽于尾侧。
继而神经板中央沿长轴下陷形成神经沟(neural groove),沟两侧边缘隆起称神经褶(neural fold),两侧神经褶在神经沟中段靠拢并愈合,愈合向两端延伸,使神经沟封闭为神经管(neural tube)。
神经管两侧的表面外胚层在管的背侧靠拢并愈合,使神经管位居于表面外胚层的深面。
神经管将分化为中枢神经系统以及松果体、神经垂体和视网膜等。
在神经褶愈合过程中,它的一些细胞迁移到神经管背侧成一条纵行细胞索,继而分裂为两条分别位于神经管的背外侧,称神经嵴(neural crest),它将分化为周围神经系统及肾上腺髓质等结构。
位于体表的表面外胚层,将分化为皮肤的表皮及其附属器,以及牙釉质、角膜上皮、晶状体、内耳膜迷路、腺垂体、口腔和鼻腔与肛门的上皮等。
2.中胚层的分化中胚层在脊索两旁从内侧向外侧依次分化为轴旁中胚层、间介中胚层和侧中胚层。
分散存在的中胚层细胞,称间充质,分化为结缔组织以及血管、肌组织等。
脊索则大部份退化消失,仅在椎间盘内残留为髓核。
(1)轴旁中胚层(paraxial mesoderm):紧邻脊索两侧的中胚层细胞迅速增殖,形成一对纵行的细胞索,即轴旁中胚层。
它随即裂为块状细胞团,称体节(somite)。
体节左右成对,从颈部向尾部依次形成,随胚龄的增长而增多,故可根据体节的数量推算早期胚龄。
第5周时,体节全部形成,共约42~44对。
体节将分化为皮肤的真皮、大部份中轴骨胳(如脊柱、肋骨)及骨骼肌。
(2)间介中胚层(intermediate mesoderm):位于轴旁中胚层与侧中胚层之间,分化为泌尿生殖系统的主要器官。
(3)侧中胚层(lateral mesoderm):是中胚层最外侧的部份,两侧的侧中胚层在口咽膜的头侧汇合为生心区。

神经系统发育了解神经系统在胚胎发育中的形成过程神经系统是人类身体中最为重要的系统之一,它负责感知和传递信息,控制身体各个部位的活动。
然而,要了解神经系统的运作机制,我们首先需要了解神经系统是如何在胚胎发育中形成的。
本文将介绍神经系统发育的过程。
一、胚胎初期神经系统的发育始于胚胎初期。
在胚胎发育的早期阶段,胚胎会分化为三个主要的胚层:内胚层、中胚层和外胚层。
神经系统的形成主要涉及内胚层和外胚层。
二、胚胎中胚层的形成在胚胎发育的过程中,内胚层会进一步分化为一个称为神经胚的结构。
神经胚是神经系统的起源,它会发展成为脊髓和大脑。
神经胚的形成是一个复杂的过程,涉及到许多细胞的分化和迁移。
三、神经胚的分化神经胚分化为两个主要的结构:中胚层和神经外胚。
中胚层发展成为脊髓,而神经外胚进一步分化为神经系统中的其他组织。
四、神经胚的迁移在神经胚形成后,胚胎会在发育过程中发生一系列的迁移。
这些迁移过程涉及到神经细胞的移动,从而形成不同的神经结构。
五、脊髓和大脑的形成神经胚中的中胚层分化为脊髓。
脊髓是神经系统的主要部分之一,负责传递感觉和运动信号。
另一方面,神经外胚的分化形成了大脑和其他神经组织。
六、突触连接的形成在神经系统发育的过程中,神经元会形成突触连接,以实现信息传递。
这些突触连接在胚胎发育中逐渐建立,并在后续的发育过程中得到进一步发展和巩固。
综上所述,神经系统在胚胎发育中的形成过程是一个复杂而精细的过程。
通过胚胎发育,内胚层和外胚层分化为神经组织,脊髓和大脑形成,并建立突触连接来传递信息。
对于理解神经系统的运作机制以及相关疾病的治疗有着重要的意义。

绪论1、发育生物学:是应用现代生物学的技术研究生物发育机制的科学。
它主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
2、(填空)发育生物学模式动物:果蝇、线虫、非洲爪蟾、斑马鱼、鸡和小鼠。
第一篇发育生物学基本原理第一章细胞命运的决定1、细胞分化:从单个的全能细胞受精卵开始产生各种分化类型细胞的发育过程称细胞分化。
2、细胞定型可分为“特化”和“决定”两个阶段:当一个细胞或者组织放在中性环境如培养皿中培养可以自主分化时,可以说这个细胞或组织发育命运已经特化;当一个细胞或组织放在胚胎另一个部位培养可以自主分化时,可以说这个细胞或组织发育命运已经决定。
(特化的发育命运是可逆的,决定的发育命运是不可逆的。
把已特化细胞或组织移植到胚胎不同部位,会分化成不同组织,把已决定细胞或组织移植到胚胎不同部位,只会分化成同一种组织。
)3、(简答)胚胎细胞发育命运的定型主要有两种作用方式:第一种通过胞质隔离实现,第二种通过胚胎诱导实现。
(1)通过胞质隔离指定细胞发育命运是指卵裂时,受精卵内特定的细胞质分离到特定的裂球中,裂球中所含有的特定胞质可以决定它发育成哪一类细胞,而及邻近细胞没有关系。
细胞发育命运的这种定型方式称为“自主特化”,细胞发育命运完全由内部细胞质组分决定。
这种以细胞自主特化为特点的胚胎发育模式称为“镶嵌型发育”,因为整体胚胎好像是由能自我分化的各部分组合而成,也称自主型发育。
(2)通过胚胎诱导指定细胞发育命运是指胚胎发育过程中,相邻细胞或组织之间通过互相作用,决定其中一方或双方细胞的分化方向。
相互作用开始前,细胞可能具有不止一种分化潜能,但是和邻近细胞或组织的相互作用逐渐限制它们的发育命运,使之只能朝一定的方向分化。
细胞发育命运的这种定型方式成为“有条件特化”或“渐进特化”或“依赖型特化”,因为细胞发育命运取决于及其邻近的细胞或组织。
这种以细胞有条件特化为特点的胚胎发育模式称为“调整型发育”,也称有条件发育或依赖型发育。
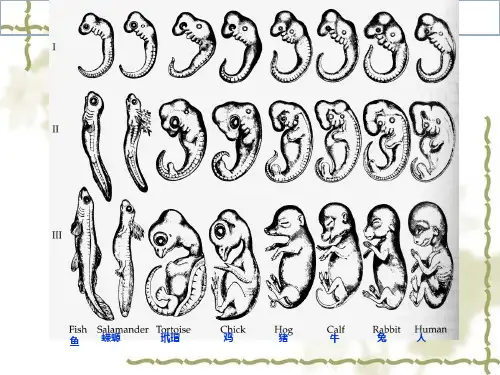
第八章神经胚和三胚层分化经过原肠作用后,胚胎已具有外、中、内三个胚层,它们是动物体所有器官形成的细胞基础。
鱼类、两栖类、爬行类、鸟类和哺乳类都具有相同的器官发生模式:外胚层形成神经系统和皮肤;内胚层形成呼吸系统和消化管;中胚层形成结缔组织、血细胞、心脏、泌尿系统以及大部分内脏器官等。
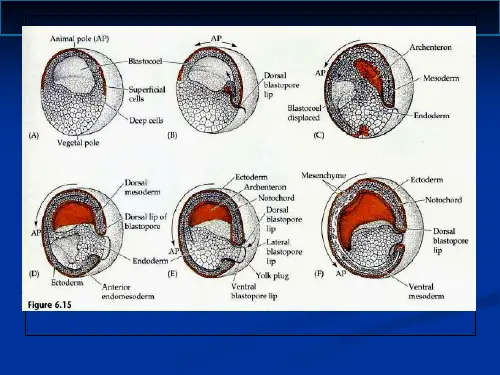
神经胚形成中胚层的分化内胚层的分化一、神经胚形成胚胎形成中枢神经系统原基即神经管的作用称为神经胚形成(neurulation),正在进行神经管形成的胚胎称为神经胚(neurula)。
神经胚形成主要由两种方式:初级神经胚形成(primary neurulation)和次级神经胚形成(secondary neurulation)。
初级神经胚形成:是指由脊索中胚层诱导覆盖于上面的外胚层细胞分裂、内陷并与表皮脱离形成中空的神经管。
次级神经胚形成:是指外胚层细胞下陷进入胚胎形成实心细胞索,接着再空洞化形成中空的神经管。
外胚层细胞的命运:背部中线区的细胞将形成脑和脊髓;中线区外侧的细胞将生成皮肤;上述二者相交处的细胞为神经嵴细胞(neural crest),它们将迁移各处形成外周神经元、色素细胞、神经胶质细胞等。
神经管形成的起始:来自背部中胚层的信号诱导预置神经板边缘的细胞的背测收缩,而预置的表皮细胞向中线移动,使表皮与神经板交接处凸起形成神经褶。
人类胚胎的神经管闭合缺陷症不同区域的神经管的封口时间不同。
第二区封口失败,胚胎的前脑不发育,即致死性的无脑症;第5区不封口导致脊柱裂口症。
Sonic Hedgehog、Pax3等因子是神经管闭合所必需的。
孕妇服用叶酸和适量的胆固醇可降低胎儿神经管缺陷的风险。
(一)、初级神经胚形成的过程:1、神经板(neural plate)形成2、神经底板(neural floor plate)形成3、神经板的整形(shaping)4、神经板弯曲成神经沟(neural groove)5、神经沟闭合形成神经管(neural tube)1. 神经板形成位于背中线处预定形成神经组织的外胚层细胞变长加厚,而预定形成表皮的细胞则变得更加扁平,使预定神经区上升到周围外胚层的上面,由此形成神经板(neural plate)。
神经板和表皮细胞都能发生固有运动(intrinsic movements)。
神经板和表皮细胞协调一致的运动最终引起神经管举起和交叠。
2. 神经底板形成以前认为只有神经板中线处细胞才能形成神经管底板,而外缘部分和神经褶则构成神经管最靠近背面的部分。
头部神经底板形成方式可能是这样的。
但最近证据表明,躯干神经底板具有独立起源,是由亨氏节一部分细胞“插入”神经板中央形成。
3. 神经板的整形和弯曲神经板的整形与神经板细胞内在力量直接有关。
神经板最主要的整形作用是通过位于脊索上面的神经板中线细胞实现的,这些细胞被称为中间铰合点。
神经板的弯曲通过神经板细胞内在力量的作用而实现,同时外胚层细胞为神经管的弯曲提供另一种动力。
4. 神经管闭合左右神经褶被牵引到背中线结合到一起,神经管随即闭合。
某些动物神经褶连接处的细胞形成神经嵴细胞,但哺乳类神经褶举起时头部神经嵴细胞就开始迁移。
神经管的形成并非在整个外胚层同时发生,如24小时鸡神经胚的尾部区域仍在进行原肠作用时,头部神经管已明显形成。
神经管前后两端的开口分别称为前端神经孔和后端神经孔。
神经管最终形成一个与表面外胚层分离的闭合的圆柱体。
神经管和表面外胚层的分离被认为是受不同的细胞粘连分子表达调节的。
神经管细胞最初都表达E-细胞选择蛋白(E-cadherin),代之以合成N-选择蛋白(N-cadherin)和N-CAM。
结果两种组织不再粘附在一起。
(二)、次级神经胚形成次级神经胚形成可以看作是原肠作用的继续,只是背唇细胞并没有内卷到胚胎内部,而是在腹面不断生长。
次级神经胚形成包括髓索形成及其随后空洞化成为神经管。
蛙和鸡胚的腰椎和尾椎形成时能观察到这种形式的神经胚形成。
特点:神经管由胚胎内细胞组成的实心索中空而成。
鸟类、哺乳类、两栖类动物胚胎的后部神经管及鱼类胚胎的全部神经管的形成采取此种方式。
(三)、脑区形成神经管同时在三个层次水平上分化成中枢神经系统:在解剖学水平,神经管及其管腔膨胀和收缩而形成脑室和脊髓的中央管。
在组织学水平,神经管壁细胞发生重排形成脑和脊髓不同的功能区域。
在细胞学水平,神经上皮细胞本身分化成身体中不同类型的神经元和神经胶质。
(四)、表皮和皮肤结构起源神经胚形成之后,覆盖胚胎表面的细胞构成预定表皮。
多数脊椎动物的表皮分为两层:外面一层为胚皮,是临时性结构,一旦底层细胞分化成表皮,胚皮便脱落;内面一层细胞称为基层或生发层,它是能形成所有表皮细胞的生发性上皮。
生发层细胞分裂产生外面另一层细胞,构成棘层。
棘层和生发层一起构成马尔皮基层。
马尔皮基层细胞再分裂产生表皮的颗粒层。
颗粒层细胞不再分裂,开始分化成表皮细胞,即角质细胞。
最终角质细胞形成角质层。
角质层细胞生成后不久就脱落,并被颗粒层新形成的细胞所取代。
有两种主要生长因子能刺激表皮发育:第一种是转化生长因子-α(transforming growth factor- α,TGF- α)。
TGF- α由基层细胞合成,并促进基层细胞自身分裂。
银屑病患者特征是大量表皮细胞脱落,可能与TGF- α过度表达有关。
另一种生长因子是角质细胞生长因子(keratinocyte growth factor, KGF),也称为成纤维细胞生长因子7(FGF7)。
KGF是真皮下面的成纤维细胞产生的一种外分泌生长因子,调节基层细胞的增殖。
中胚层的分化脊椎动物中胚层的分化对于器官和系统的发生起着主导和奠基的作用。
其中脊索是这一阶段发育的启动和组织者,而在脊索和神经管的作用下,中胚层分化深入。
脊椎动物中胚层的分化发育与神经胚的形成几乎是同时进行,相互促进的,而神经胚发育的完成又为中胚层的进一步发育奠定了形态结构以及诱导控制环境的基础。
中胚层对机体结构与机能进一步发展的意义:由于中胚层的形成减轻了内、外胚层的负担,引起了一系列组织、器官、系统的分化,为动物体结构的进一步复杂完备提供了必要的物质条件。
由于中胚层的形成,促进了新陈代谢的加强。
中胚层的形成也是动物由水生进化到陆生的基本条件之一。
(一)、神经胚时期中胚层可分成5个区域:1. 脊索中胚层(chordamesoderm)形成脊索;2. 背部体壁中胚层(somitic dorsal mesoderm)形成体节和神经管两侧的中胚层细胞;3. 中段中胚层(intermediate mesoderm)形成泌尿系统和生殖器官;4. 侧板中胚层(lateral plate )形成心脏、血管、血细胞;5. 头部间质(head mesenchyme)形成面部结缔组织和肌肉。
二)、心脏的发生形成循环系统是侧板中胚层最重要的功能之一。
循环系统是发育过程中最先行使功能的系统,而心脏是第一个行使功能的器官。
脊椎动物的心脏起源于脏壁中胚层的两个区域(心脏原基),分别位于身体的两侧。
这两个区域与周围临近组织的相互作用决定心脏发育。
三)、血管的形成血管形成是生理限制、物理限制和进化限制协调统一的产物。
由中胚层形成血管称为血管形成。
鸡胚有两种成血管细胞。
第一种是脑区轴旁中胚层提供形成头部血管的成血管细胞。
第二种成血管细胞来自脏壁中胚层,它们移居到内脏器官、肠和主动脉基底部。
心脏发育的主要阶段:预定心脏形成区的形成(预定心脏形成细胞通过原条迁移形成心脏原基)。
心脏形成细胞的分化。
两个心脏原基合拢形成心管。
心脏腔室的分隔,心脏四腔形成。
有三种生长因子可能参与血管形成的启动:第一种是碱性成纤维细胞生长因子(FGF2),为中胚层细胞形成成血管细胞所必需。
第二种是血管内皮生长因子(VEGF),VEGF能特异性地促进成血管细胞分化,并促进成血管细胞分裂增殖,形成血管内皮。
第三种是血管生成素angiopoietin-1,能调节内皮细胞和平滑肌之间的相互作用。
(四)、血细胞的发育干细胞(stem cell)是能够保持胚胎细胞特性,大量增殖以产生更多的干细胞和更多的分化细胞的细胞群,它们在成年体内连续不断地经历进一步的发育变化。
有些干细胞锁定在G0期,而另一些干细胞则处于活跃的细胞周期中。
干细胞后代进入的发育途径依赖于它所处位置的分子环境。
干细胞存在于所有组织中,最容易研究且研究最多的干细胞是多能造血干细胞的发育。
多能造血干细胞可以产生所有类型的血细胞,Till和McCulloch(1961)证实了它们的存在。
最早的多能造血干细胞是CFU-M,L。
CFU-M,L的发育取决于转录因子SCL。
缺少SCL的小鼠将死于血细胞及淋巴细胞缺乏。
CFU-M,L能产生CFU-S细胞(血细胞)和CFU-L细胞(淋巴细胞),两者都是多能干细胞,它们的后代能分化成大量不同类型的细胞。
CFU-S形成的直接后代是限制系干细胞,每个细胞除更新自己外只能产生一种类型的细胞。
有些造血生长因子(如IL-3)可以刺激较原始干细胞的分裂和成熟,使所有类型的血细胞数量增加。
另一些生长因子只能刺激特定的细胞系细胞分裂和成熟。
细胞对生长因子的反应能力取决于细胞表面是否存在生长因子受体。