植物生理学:第三节 呼吸链和氧化磷酸化
- 格式:ppt
- 大小:7.10 MB
- 文档页数:22

有氧呼吸名词解释植物生理学有氧呼吸是植物生理学中一个重要的过程,它指的是植物细胞中利用氧气(O2)氧化有机物质以产生能量的过程。
有氧呼吸是植物细胞中能量供应的关键途径之一,通过此过程,植物能够从有机物中释放出能量,并将其用于维持生命活动、生长和发育。
以下是有关有氧呼吸的详细解释,涵盖了其机制、调控和重要性。
一、有氧呼吸机制:有氧呼吸是一个复杂的过程,可以分为三个主要阶段:糖解、三羧酸循环和氧化磷酸化。
下面对这些阶段进行详细解释:1. 糖解(Glycolysis):糖解是有氧呼吸的起始阶段,发生在细胞质中。
在糖解过程中,葡萄糖分子被分解成两个三碳化合物,即丙酮酸和磷酸二酸(pyruvate)。
这个过程产生少量的ATP(三磷酸腺苷)和NADH(还原型辅酶NAD+),同时释放出一些水和热能。
2. 三羧酸循环(Citric Acid Cycle):三羧酸循环也被称为克雷布循环或柠檬酸循环,它是有氧呼吸的第二个阶段,发生在线粒体的基质中。
在三羧酸循环中,每个磷酸二酸分子被氧化成二氧化碳,并释放出高能电子,这些电子被捕获到辅酶NAD+和辅酶FAD(腺嘌呤核苷二核苷酸)中,生成NADH和FADH2。
同时,三羧酸循环还产生一些ATP和GTP(三磷酸鸟苷)。
3. 氧化磷酸化(Oxidative Phosphorylation):氧化磷酸化是有氧呼吸的最后一个阶段,也是能量产生的主要阶段。
它发生在线粒体的内膜上的呼吸链中。
在氧化磷酸化过程中,NADH和FADH2释放出的高能电子被逐步通过一系列的酶和辅酶传递,最终被氧气接受,生成水。
这个过程中产生的能量被用于推动ATP合成酶,使其合成ATP。
这个过程称为化学耦合,因为能量释放与ATP 合成紧密耦合在一起。
二、有氧呼吸的调控:植物细胞中的有氧呼吸受到多种内外因素的调控。
以下是一些常见的调控因素:1. 氧气浓度:有氧呼吸需要氧气作为底物,因此氧气浓度是一个重要的调控因素。

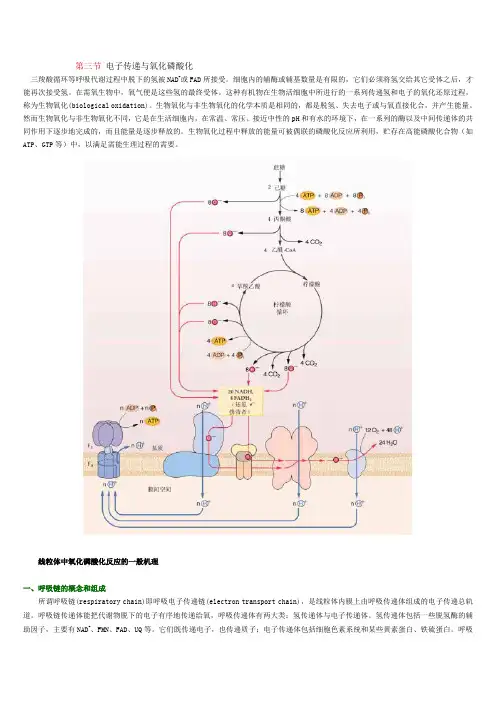
第三节电子传递与氧化磷酸化三羧酸循环等呼吸代谢过程中脱下的氢被NAD+或FAD所接受。
细胞内的辅酶或辅基数量是有限的,它们必须将氢交给其它受体之后,才能再次接受氢。
在需氧生物中,氧气便是这些氢的最终受体。
这种有机物在生物活细胞中所进行的一系列传递氢和电子的氧化还原过程,称为生物氧化(biological oxidation)。
生物氧化与非生物氧化的化学本质是相同的,都是脱氢、失去电子或与氧直接化合,并产生能量。
然而生物氧化与非生物氧化不同,它是在生活细胞内,在常温、常压、接近中性的pH和有水的环境下,在一系列的酶以及中间传递体的共同作用下逐步地完成的,而且能量是逐步释放的。
生物氧化过程中释放的能量可被偶联的磷酸化反应所利用,贮存在高能磷酸化合物(如ATP、GTP等)中,以满足需能生理过程的需要。
线粒体中氧化磷酸化反应的一般机理一、呼吸链的概念和组成所谓呼吸链(respiratory chain)即呼吸电子传递链(electron transport chain),是线粒体内膜上由呼吸传递体组成的电子传递总轨道。
呼吸链传递体能把代谢物脱下的电子有序地传递给氧,呼吸传递体有两大类:氢传递体与电子传递体。
氢传递体包括一些脱氢酶的辅助因子,主要有NAD+、FMN、FAD、UQ等。
它们既传递电子,也传递质子;电子传递体包括细胞色素系统和某些黄素蛋白、铁硫蛋白。
呼吸链传递体传递电子的顺序是:代谢物→NAD+→FAD→UQ→细胞色素系统→O2。
呼吸链中五种酶复合体(enzyme complex)的组成结构和功能简要介绍如下(图5-11,5-12)。
图 5-11 植物线粒体内膜上的复合体及其电子传递Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ分别代表复合体Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ; UQ库代表存在于线粒体中的泛醌库1.复合体Ⅰ 又称NADH∶泛醌氧化还原酶(NADH∶ubiquinone oxidoreductase)。
分子量700X103~900X103,含有25种不同的蛋白质,包括以黄素单核苷酸(flav in mononucleotide,FMN)为辅基的黄素蛋白和多种铁硫蛋白,如水溶性的铁硫蛋白(iron sulfur protein,IP)、铁硫黄素蛋白(iron sulfur flavoprotein,FP)、泛醌(ubiquinone,UQ)、磷脂(phospholipid)。

植物生理学名词解释1、渗透势:由于溶质作用使细胞水势降低的值。
2 呼吸商:植物在一定时间内放出的CO2与吸收O2的比值。
3 荧光现象:叶绿素吸收的光能从第一单线态以红光的形式散失,回到基态的现象。
4 光补偿点:光饱和点以下,使光合作用吸收的CO2与呼吸作用放出的CO2相等的光强。
6 生长调节剂:人工合成的,与激素功能类似,可调节植物生长发育的活性物质。
7 生长:由于细胞分裂和扩大引起的植物体积和重量的不可逆增加。
8 光周期现象:植物通过感受昼夜长短的变化而控制开花的现象。
9 逆境:对植物生长发育有利的各种环境因素的总称。
10 自由水:在植物体内不被吸附,可以自由移动的水。
11、C02补偿点:植物光合同化C02量与呼吸释放C02量达到动态平衡时,环境C02含量。
12. 植物细胞全能性:植物的每个细胞均含有母体的全套基因,并在适宜条件下均能发育成完整个体的潜在能力。
13、氧化磷酸化:是指电子通过呼吸链传递给分子氧和生成水,并偶联ADP和磷酸生成A TP的过程。
14、源-库单位:代谢源与代谢库及其二者之间的输导组织;或同化物供求上有对应关系的源与库的合称。
15.乙烯的三重反应:随着浓度的升高,乙烯抑制茎的伸长生长、促进茎或根的横向增粗以及茎的横向地性生长的现象。
16、P680:光合作用中光系统II(PSII)的中心色素分子,主要特征是吸收680nm的红光,并进行光化学反应。
17、PEP:磷酸烯醇式丙酮酸,为C4循环途径中C02的受体,与C02结合形成草酰乙酸。
18.RQ:为呼吸商,指植物呼吸过程中,放出的体积与吸收O的体积之比。
19.逆境蛋白:逆境环境,如干旱、高温、低温、盐碱、病原菌、紫外线等诱导植物体内形成新的蛋白质的统称。
20诱导酶又叫适应酶。
指植物体内本来不含有,但在特定外来物质的诱导下可以生成的酶。
21、光饱和点:在光照强度较低时,光合速率随光强的增加而相应增加;光强进一步提高时,光合速率的增加逐渐减小,当超过一定光强时即不再增加,这种现象称光饱和现象。

基本内容第三节电子传递与氧化磷酸化(electron transport and oxidative phosphorylation)。
有机物质在生物体细胞内进行氧化分解,生成二氧化碳、水和释放能量的称为生物氧化(biological oxidation)。
一、呼吸链(respiratory chain)糖酵解和三羧酸循环中所产生的NADH+H+不能直接与游离的氧分子结合,需要经过电子传递链传递后,才能与氧结合。
电子传递链(electron transport chain)亦称呼吸链(respiratory chain),就是呼吸代谢中间产物的电子和质子,沿着一系列有顺序的电子传递体组成的电子传递途径,传递到分子氧的总过程。
组成电子传递链的传递体可分为氢传递体和电子传递体。
氢传递体传递氢(包括质子和电子,以2H++2e-表示),它们作为脱氢酶的辅助因子,有下列几种:NAD(即辅酶Ⅰ)、NADP(即辅酶Ⅱ)、黄素单核苷酸(FMN)和黄素腺嘌呤二核苷酸(FAD),它们都能进行氧化还原反应。
电子传递体是指细胞色素体系和铁硫蛋白(Fe-S),它们只传递电子。
细胞色素是一类以铁卟啉为辅基的结合蛋白质,根据吸收光谱的不同分为a、b和c 3类,每类又再分为若干种。
细胞色素传递电子的机理,主要是通过铁卟啉辅基中的铁离子完成的,Fe3+在接受电子后还原为Fe2+,Fe2+传出电子后又氧化为Fe3+。
植物线粒体的电子传递链位于线粒体的内膜上,由5种蛋白复合体(protein complex)组成(图4-6)。
1、复合体Ⅰ(complex I)也称NADH脱氢酶(NADH dehydrogenase),由结合紧密的辅因子FMN和几个Fe-S中心组成,其作用是将线粒体基质中的NADH+H+的2对电子即4个质子泵到膜间间隙(intermembrane space),同时复合体也经过Fe-S中心将电子转移给泛醌(ubiquinone, UQ或Q)。

氧化磷酸化名词解释生物化学一、氧化磷酸化名词解释呼吸链的主要功能是产生能量货币ATP。
当电子沿着呼吸链向下游传递的时候总伴随着自由能的释放,释放的自由能有很大一部分用来驱动ATP的合成,这种与电子传递偶联在一起的合成ATP方式被称为氧化磷酸化(OxP)。
二、氧化磷酸化的偶联机制1、化学渗透学说该学说由Peter Mitchell于1961年提出,其核心内容是电子在沿着呼吸链向下游传递的时候,释放的自由能转化为跨线粒体内膜(或跨细菌质膜)的质子梯度,质子梯度中蕴藏的电化学势能直接用来驱动ATP的合成。
驱动ATP合成的质子梯度通常被称为质子驱动力(pmf),它由化学势能(质子的浓度差)和电势能(内负外正)两部分组成。
支持化学渗透学说的主要证据:•氧化磷酸化的进行需要完整的线粒体内膜的存在。
•使用精确的pH计可以检测到跨线粒体内膜的质子梯度存在。
据测定,一个呼吸活跃的线粒体的膜间隙的pH要比其基质的pH 低0.75个单位。
•破坏质子驱动力的化学试剂能够抑制ATP的合成。
•从线粒体内膜纯化得到一种酶能够直接利用质子梯度合成ATP,此酶称为F1F0-ATP合酶。
•人工建立的跨线粒体内膜的质子梯度也可驱动ATP的合成2、结合变化学说1977年Paul D. Boyer提出的结合变化学说能正确地解释F1F0-ATP 合酶的作用机理。
结合变化学说可简化为:质子流动→驱动C单位转动→带动γ亚基转动→诱导β亚基构象变化→ATP释放和重新合成。
支持结合变化学说的证据:•18O同位素交换实验•John Walker获得的F1的晶体结构清楚地表明,3个β亚基处于不同的构象并和不同的核苷酸配体结合•日本科学家采取特别的手段直接观察到F1的旋转催化三、氧化磷酸化的解偶联氧化磷酸化与呼吸链通常是紧密偶联的,但是,低水平的质子泄漏时刻发生在线粒体内膜上,因此,确切地说,线粒体通常是部分解偶联的。
解偶联一般是受解偶联剂作用所致。
解偶联剂的作用机制在于它们能够快速地消耗跨膜的质子梯度,使得质子难以通过F1F0-ATP合酶上的质子通道来合成ATP,从而将贮存在质子梯度之中的电化学势能转变成热。

第三章植物的呼吸作用一、名词解释1.呼吸作用2.有氧呼吸3.无氧呼吸4.呼吸跃变5.氧化磷酸化6.P/O 7.无氧呼吸消失点8.抗氰呼吸9.糖酵解10.三羧酸循环11.磷酸戊糖途径12.巴斯德效应13.末端氧化酶14.温度系数15.呼吸链16.氧化磷酸化17.伤呼吸18.呼吸效率19.呼吸商20.呼吸速率.二、写出下列符号的中文名称1. EMP2. FAD3. DHAP4. GSSC5. FMA6. GAC7.Cyt8. GAS9. HMP 10. FP11. UQ 12. PPP 13. TCA14.GAP 15. DNP三、填空题1.呼吸作用的糖酵解是在细胞的进行的,磷酸戊糖途径是在细胞的进行,三羧酸循环是在进行。
2.有氧呼吸和无氧呼吸的主要区别是______,它们开始走的共同途径是______。
3. 植物组织衰老时,磷酸戊糖支路在呼吸代谢途径中所占比例。
4.一分子的葡萄糖经过糖酵解净产生______个A TP。
5. 植物呼吸代谢多样性表现在、和。
6. 糖酵解和戊糖磷酸途径之间有一个重要区别,即氧化还原的辅酶不同,糖酵解是______,而戊糖磷酸途径是______。
7.呼吸商为1.0时,说明被氧化为,呼吸商<1.0时,被氧化物为,大于1.0时,被氧化物为。
8. 调节控制糖酵解过程的反应速度时,催化三个主要控制反应的酶是______、______、______。
糖酵解的酶系定位于______内,三羧酸循环酶系定位于______内,呼吸链的组分定位于______。
9. 酚氧化酶是一种含______的氧化酶,存在于______,______内。
这种酶在制茶中有重要作用,在制绿茶时要立即杀青,防止______,避免______产生,保持茶气清香。
10. 产生丙酮酸的糖酵解过程是______与______的共同途径。
11. 无氧呼吸的特征是______,底物氧化降解______,大部分底物仍是______,因而释放______少。

呼吸链与氧化磷酸化呼吸链是细胞内的一个重要代谢过程,负责产生细胞所需的能量。
而氧化磷酸化是呼吸链中的最后一步,它将呼吸链过程中所产生的能量转化为细胞内的三磷酸腺苷(ATP)。
本文将详细介绍呼吸链和氧化磷酸化的相关知识。
一、呼吸链的概述呼吸链是细胞内的一系列酶和分子复合物组成的蛋白质链,其作用是将有机物质在细胞内氧化分解,同时释放出能量。
呼吸链的过程可以分为四个复合物:复合物Ⅰ、复合物Ⅱ、复合物Ⅲ和复合物Ⅳ,以及两个载体分子:辅酶Q和细胞色素C。
1. 复合物Ⅰ:复合物Ⅰ是呼吸链的起始点,它将NADH的电子和质子转移到辅酶Q上,并释放出能量。
2. 复合物Ⅱ:复合物Ⅱ接受来自有机物质的电子,并将它们转移到辅酶Q上。
3. 复合物Ⅲ:复合物Ⅲ将辅酶Q上的电子和质子转移到细胞色素C 上,细胞色素C则将电子传递给复合物Ⅳ。
4. 复合物Ⅳ:复合物Ⅳ将电子从细胞色素C转移到最终受体氧气上,同时将质子跨膜输运,形成质子梯度。
5. 辅酶Q和细胞色素C:辅酶Q和细胞色素C作为电子和质子的载体,在呼吸链中不断传递电子,并参与质子泵运输。
二、氧化磷酸化的概述氧化磷酸化是呼吸链的最后一步,其作用是将呼吸链过程中产生的质子梯度和电子传递的能量转化为细胞内的三磷酸腺苷(ATP)。
氧化磷酸化包括两个主要过程:氧子耦联和ATP合成。
1. 氧子耦联:氧子耦联是通过复合物Ⅳ将电子传递给氧气,形成水的过程。
在这个过程中,复合物Ⅳ释放出能量,用来推动质子向细胞外膜跨越,并形成质子梯度。
2. ATP合成:ATP合成是质子从细胞外膜返回细胞内膜的过程,通过ATP合酶(ATP synthase)将ADP和磷酸基团结合生成ATP。
这个过程被称为化学耦联,因为它将质子梯度的能量转化为ATP的化学能。
三、呼吸链和氧化磷酸化的相互关系呼吸链和氧化磷酸化是密切相关的,呼吸链产生的质子梯度提供了氧化磷酸化所需的能量。
而氧化磷酸化则利用质子梯度推动ATP的合成,为细胞提供能量。

氧化磷酸化名词解释植物生理学
氧化磷酸化,生物化学过程,在真核细胞的线粒体或细菌中,是物质在体内氧化时释放的能量通过呼吸链供给ADP与无机磷合成ATP的偶联反应。
作用:
氧化磷酸化作用是指有机物包括糖、脂、氨基酸等在分解过程中的氧化步骤所释放的能量,驱动ATP合成的过程。
在真核细胞中,氧化磷酸化作用在线粒体中发生,参与氧化及磷酸化的体系以复合体的形式分布在线粒体的内膜上,构成呼吸链,也称电子传递链。
其功能是进行电子传递、H+传递及氧的利用,产生H2O和ATP。
扩展:这种复合体一般有四个部分组成:复合体1.NADH-Q还原酶,复合体2.琥珀酸—Q还原酶.复合体3.细胞色素还原酶.4细胞色素氧化酶。
电子在电子载体的传递过程为:NADH或FADH2 --Q(泛醌)——细胞色素c——O2(形成水和ATP的过程)。


第四章呼吸作用一、名词解释1、呼吸作用:生物体内的有机物质通过氧化还原而产生CO2,同时释放能量的过程。
2、有氧呼吸:指生活细胞在氧气的参与下,把某些有机物质彻底氧化分解,放出CO2并形成水,同时释放能量的过程。
3、三羧酸循环:丙酮酸在有氧条件下由细胞质进入线粒体逐步氧化分解,最终生成水和二氧化碳。
4、生物氧化:指有机物质在生物体内进行氧化分解,生成CO2和H2O,放出能量的过程。
5、呼吸链:呼吸代谢中间产物的电子和质子,沿着一系列有序的电子传递体组成的电子传递途径,传递到氧分子的总轨道。
6、氧化磷酸化:在生物氧化过程中,电子经过线粒体的呼吸链传递给氧(形成水分子),同时使ADP被磷酸化为ATP的过程。
7、呼吸商:又称呼吸系数。
是指在一定时间内,植物组织释放CO2的摩尔数与吸收氧的摩尔数之比。
8.糖酵解:胞质溶胶中的己糖在无氧或有氧状态下分解成丙酮酸的过程。
二、填空题1、呼吸作用的糖的分解代谢途径中,糖酵解和戊糖磷酸途径在细胞质中进行;三羧酸循环途径在线粒体中进行。
三羧酸循环是英国生物化学家Krebs 首先发现的。
2、早稻浸种催芽时,用温水淋种和时常翻种,其目的就是使呼吸作用正常进行。
当植物组织受伤时,其呼吸速率加快。
春天如果温度过低,就会导致秧苗发烂,这是因为低温破坏了线粒体的结构,呼吸“空转”,缺乏能量,引起代谢紊乱的缘故。
3.呼吸链的最终电子受体是O2氧化磷酸化与电子传递链结偶联,将影响_ ATP _的产生。
4.糖酵解是在细胞细胞基质中进行的,它是有氧呼吸和无氧呼吸呼吸的共同途径。
5.氧化磷酸化的进行与ATP合酶密切相关,氧化磷酸化与电子传递链解偶联将影响__ ATP__的产生。
6.植物呼吸过程中,EMP的酶系位于细胞的细胞基质部分,TCA的酶系位于线粒体的线粒体基质部位,呼吸链的酶系位于线粒体的嵴部位。
7. 一分子葡萄糖经有氧呼吸彻底氧化,可净产生__38__分子ATP,•需要经过__6_底物水平的磷酸化。
植物生理学复习1.氧化磷酸化:是指呼吸链上的氧化过程,伴随着ADP被磷酸化为ATP的作用。
2.P/O:指呼吸链中每消耗1个氧原子与用去Pi或产生ATP的分子数。
3.抗氰呼吸:某些植物组织对氰化物不敏感的那部分呼吸。
即在有氰化物存在的情况下仍能够进行其它的呼吸途径。
4.糖酵解:指在细胞质内所发生的,由葡萄糖分解为丙酮酸的过程。
5.三羧酸循环:丙酮酸在有氧条件下,通过一个包括三羧酸和二羧酸的循环而逐步氧化分解CO2的过程。
6.末端氧化酶:是指处于生物氧化作用一系列反应的最末端,将底物脱下的氢或电子传递给氧,并形成H2O或H2O2的氧化酶类。
7.呼吸链:呼吸代谢中间产物随电子和质子,沿着一系列有顺序的电子传递体组成的电子传递途径,传递到分子氧的总轨道。
8.氧化磷酸化:是指呼吸链上的氧化过程,伴随着ADP被磷酸化为ATP的作用。
9.植物生长物质:调节植物生长发育的物质。
10.植物激素:是指一些在植物体内合成,并从产生之处运往作用部位,对生长发育起调控作用的微量(0.5)有机物。
11.植物生长调节剂:指一些具有植物激素活性的人工合成的物质。
12.三重反应:乙烯可抑制黄化豌豆幼苗上胚轴的伸长生长,促进其加粗生长,地上部分失去负向地性生长(偏上生长)。
13.极性运输: 是指生长素只能从植物体的形态学上端向下端运输。
14.程序性细胞死亡:是指为维持内环境稳定,由基因控制的细胞自主的有序性的死亡,它涉及一系列基因的激活、表达以及调控等的作用,因而是具有生理性和选择性的。
15.细胞全能性:植物的每个细胞均含有母体的全套基因,并在适宜条件下均能发育成完整个体的潜在能力。
16.愈伤组织: 原是指植物在受伤后于伤口表面形成的一团薄壁细胞,在组培中,则指在人工培养基上由外植体长出来的一团无序生长的薄壁细胞。
特征:细胞排列疏松而无规则,是一种高度液泡化的呈无定形状态的薄壁细胞.17.脱分化: 是指分化细胞失去特有的结构和功能变为未分化细胞特性的过程,即分化的细胞在适当的条件下转变为胚性状态而重新获得分裂能力的过程。
《植物生理学》第三章植物的呼吸作用复习题及答案一、名词解释1.呼吸作用(respiration):生活细胞内的有机物,在酶的参与下,逐步氧化分解并释放能量的过程。
2.有氧呼吸(aerobic respiration):生活细胞利用分子氧,将某些有机物质彻底氧化分解,形成CO2和H2O,同时释放能量的过程。
3.无氧呼吸(anaerobic respiration):生活细胞在无氧条件下,把某些有机物分解成为不彻底的氧化产物,同时释放能量的过程。
微生物的无氧呼吸通常称为发酵(fermentation)。
4. 呼吸跃变(respiratory climacteric):果实成熟过程中,呼吸速率突然增高,然后又迅速下降的现象。
呼吸跃变的产生与外界温度和果实内乙烯的释放密切相关。
呼吸跃变是果实进入完熟的一种特征,在果实贮藏和运输中,重要的问题是降低温度,抑制果实中乙烯的产生,推迟呼吸跃变的发生,降低其发生的强度,延迟果实的完熟。
5. 氧化磷酸化(oxidative phosphorylation):在线粒体内膜上电子经电子传递链传递给分子氧生成水,并偶联ADP和Pi生成ATP的过程。
它是需氧生物生物氧化生成ATP的主要方式。
6. P/O:磷氧比,氧化磷酸化的活力指标,指每吸收一个氧原子所能酯化的无机磷的数目,即有几个无机磷与ADP形成了ATP。
呼吸链中两个质子和两个电子从NADH+H+开始传至氧生成水,一般可形成3分子的ATP,其P/O比为3。
7. 无氧呼吸消失点(anaerobic respiration extinction point):无氧呼吸停止进行的最低氧浓度(10%左右)称为无氧呼吸消失点。
8.抗氰呼吸(cyanide resistant respiration) :对氰化物不敏感的那一部分呼吸。
抗氰呼吸可以在某些条件下与电子传递主路交替运行,因此,这一呼吸支路又称为交替途径(alternative pathway)。
氧化磷酸化和呼吸链复合体
氧化磷酸化是细胞内能量产生的主要途径之一,它通过将ADP和Pi反应生成ATP来提供细胞所需的能量。
而呼吸链复合体则是实现氧化磷酸化的关键组成部分。
氧化磷酸化和呼吸链复合体之间的关系紧密相连。
氧化磷酸化的过程需要通过呼吸链复合体中的一系列酶的协同作用来完成,其中涉及到许多生物大分子的参与。
这些生物大分子中包括多种酶类、蛋白质、脂类和核酸等。
氧化磷酸化的过程涉及到三个蛋白复合体:NADH-脱氢酶复合体、细胞色素c氧化酶复合体和ATP合成酶复合体。
其中,NADH-脱氢酶复合体和细胞色素c氧化酶复合体是呼吸链复合体中的两个关键组
成部分。
它们通过不同的途径将氢离子从细胞内转移到细胞外,从而产生电化学梯度,为ATP的合成提供动力。
总之,氧化磷酸化和呼吸链复合体是细胞内多个生物大分子的协同作用,实现了细胞内的能量转化和供能。
对于细胞的正常运转和生命活动具有非常重要的意义。
- 1 -。